ландии,
годовой энергобаланс характеризовался
следующими пара-
метрами
(Дж/м2):
потребление пищи 473, рост 210, дыхание
750,
фекалии
184, потеря с метаболитами 193 (МП1е1, Мапп,
1973). В од-
ном
из озер Англии популяция водяного
ослика со средней биомас-
сой
752 мг/м2
потребляет в год на 1 м2
570 кДж, ассимилирует
231
кДж, тратит на дыхание 92 кДж и аккумулирует
39 кДж (Ай-
соск,
1982). На примере энергобаланса морских
ежей видно, как
велика
доля энергии выделяемых метаболитов,
которая во многих
работах
не учитывается. У личинок хирономид
энергия метаболитов
равна
10—15% энергии рациона, у литорального
моллюска Те^и1а
[шеЬгаНз
потеря энергии с метаболитами в 1,5 раза
выше энергии
прироста.
Величина
рассеяния энергии популяцией, с одной
стороны, про-
порциональна
ее биомассе, а с другой — интенсивности
метаболиз-
ма,
свойственной особям. Поскольку с
уменьшением размеров ор-
ганизмов
интенсивность их метаболизма повышается,
из двух по-
пуляций
с равной биомассой рассеяние энергии
будет больше в
той,
которая состоит из мелких особей. Точно
так же следует оце-
нивать
и роль молоди в популяциях крупных
организмов. Помимо
размерного
признака, не следует забывать и о
видовых различиях
уровня
метаболизма. При прочих равных условиях
популяции, со-
стоящей
из особей с высоким уровнем обмена,
для поддержания
существования
требуется больше энергии, чем популяции,
образо-
ванной
организмами с низкой интенсивностью
метаболизма.
Знание
отдельных параметров энергобаланса
и баланса веществ
позволяет
правильно оценить роль популяций
гидробионтов в эко-
системных
процессах и перспективность их
хозяйственного освое-
ния,
в частности при организации аквакультуры.
ВОСПРОИЗВОДСТВО
И ДИНАМИКА ПОПУЛЯЦИИ
ГИДРОБИОНТОВ
Каждая
популяция представляет собой систему
с очень лабиль-
ными
параметрами; они обеспечивают такие
варианты ее состоя-
ния,
которые оптимальны в каждый момент
времени применитель-
но
к меняющимся условиям существования.
К последним относятся
не
только абиотические воздействия, но
и все другие, включая пе-
ремены
во внутрисистемных взаимодействиях.
Как организм в от-
вет
на те или иные сигналы перестраивает
структуру и функции
(увеличение
дыхательных поверхностей при дефиците
кислорода,
изменение
уровня обмена во время роста), так
популяция являет
собой
цепь разных состояний, каждое из которых
— оптимальный
вариант
адаптивного ответа (соответствующий
уровню «вычисли-
тельных
возможностей» данной популяции). Эти
оптимумы состоя-
ний
ориентированы в основном на поддержание
организованности
системы
в условиях дезорганизующих воздействий
среды и на мак-
9*
259Глава 9
симальное
использование имеющихся возможностей
ввода энергии
в
систему за счет наиболее полной
утилизации жизненных ресур-
сов.
Чем
больше вводится энергии в популяцию,
тем больше возмож-
ность
повышения ее организованности как
системы. Другими сло-
вами,
увеличение численности особей и
интенсификация их функ-
ций
— важнейшее средство повышения
организации популяций и
залог
расширения их адаптивных возможностей.
Однако на этом
пути
есть и ограничение: популяция — один
из элементов более
сложной
системы — биоценоза, и ее численность
должна быть
согласована
с его структурой. Если, например,
количество альго-
фагов
и их корма — водорослей — окажется
несогласованным,
число
и рацион потребителей должны прийти
в соответствие с
имеющимися
кормовыми ресурсами. Если рассмотреть
явление в
более
широком плане, то не только уничтожение
запасов корма,
но
и все другие воздействия популяции на
среду, сопровождаю-
щиеся
ее ухудшением, не должны выходить за
предел тех «репа-
раций»,
которые в состоянии обеспечить биоценоз.
Важнейшие
параметры популяции — ее величина и
ее функцио-
нальная
активность, которая также определяется
главным образом
количеством
особей. Оба эти параметра в основном
регулируются,
поддерживаясь
на оптимальном уровне, сопряженными
друг с дру-
гом
процессами — рождаемостью
и смертностью.
Сдвиги в соотно-
шении
скорости этих процессов обусловливают
динамику числен-
ности
и биомассы популяции, тип которой
специфичен для каждого
вида
и, как всякий видовой признак, имеет
адаптивный характер.
Из
этого положения автоматически вытекает,
что рождаемость и
смертность
сами являются регулируемыми свойствами
популяции,
хотя
диапазон и особенности их изменения
в значительной мере
закреплены
наследственно.
Рождаемость
Пополнение
популяции новыми особями может
выражаться
в
абсолютных и относительных показателях.
Различают рождае-
мость
за тот или иной срок (АЛО, за единицу
времени (АЛ^/А<) и
уровень
рождаемости (АЫ/ЫА(),
т. е. величину пополнения в еди-
ницу
времени, отнесенную к числу особей в
исходной популяции.
Рождаемость
за единицу времени или мгновенную
скорость вос-
произведения
для популяций многих животных вычисляют
по фор-
муле
(Ейтопзоп, 1968): У=кп[1,
где V
— показатель рождаемости
(АN);
п
— число самок в популяции, к
— среднее число яиц на
каждую
самку с яйцами, I
— длительность развития яиц в сутках.
Необходимый
уровень рождаемости в популяции
обеспечивается
маневренным
использованием разных форм и ритмов
размножения,
а
также регулированием плодовитости
особей.
Формы
размножения. Для каждой популяции
характерна та или
иная
форма размножения, причем в ряде случаев
она зависит от
внешних
условий. Это наблюдается, например,
при чередовании по-
260
лового
и бесполого размножения, гетерогонии,
метагенезе, смене
гермафродитизма
и раздельнополости. Смена полового и
бесполого
размножения
в их типичной форме (цитогония)
наблюдается в по-
пуляциях
многих простейших и водорослей. Переход
к половому
размножению
обычно наступает с ухудшением условий
существо-
вания
и ведет к снижению рождаемости, вместе
с тем повышая
жизнестойкость
пополнения. Метагенез свойствен многим
кишечно-
полостным.
Вегетативное размножение у губок,
мшанок, асцидий
и
других животных повышает эффективность
пополнения популя-
ций
в неблагоприятных условиях, когда
образующиеся стойкие ста-
дии
(геммулы губок, статобласты мшанок)
обеспечивают воспро-
изводство
популяции, прерванное на тот или иной
период (высы-
хание
и промерзание водоемов). Гетерогония
чрезвычайно широко
распространена
в популяциях низших ракообразных и
коловраток.
Нередко
форма размножения определяется
условиями освеще-
ния.
Рачки ЬаЫсосега
аезНьа
в зависимости от длины светового
дня
образуют субитанные или диапаузирующие
яйца (Магсиз,
1982),
коловратки Ыо1отта1а
сореиз
в темноте размножаются толь-
ко
партеногенетически и преимущественно
двуполо при длинном
световом
дне (Роигпо! е! а11., 1981). У ряда коловраток
переход к
двуполому
размножению стимулирует токоферол,
присутствующий
в
пище, который вырабатывается только
растениями. Преимущест-
ва,
которые дает партеногенез, особенно
важны в местообитаниях
с
нестабильным режимом (пересыхание
водоемов и др.).
От
условий существования популяции, в
первую очередь пище-
вых
и температурных, зависит соотношение
между числом парте-
ногенетических
и двуполых поколений. Различают виды
моноцик-
лические
— с одним поколением самцов в году,
дициклические
—
с
двумя, полициклические
— со многими и ациклические
— размно-
жающиеся
почти исключительно партеногенетически.
В мелких,
сильно
прогреваемых лужах число двуполых
поколений может
быть
больше, чем в озерах. Так, в средних
широтах рачок 8то-
серНа1и$,
обитающий в лужах, дицикличен, а в
озерах — моноцик-
личен.
Многие ОарЪтйае, на Крайнем Севере
моноциклические или
даже
ациклические, с продвижением к югу
становятся сначала ди-
циклическими,
а затем и полициклическими.
Ритмы
размножения. Обычно размножение в
популяциях гидро-
бионтов
происходит с различной интенсивностью
во времени и при-
урочено
к тем или иным периодам, в результате
чего возникает
определенный
ритм размножения. Его существование
объясняется,
с
одной стороны, необходимостью
синхронизации образования но,-
вых
поколений с наступлением условий для
роста и развития мо-
лоди;
с другой стороны, вымет гамет будет-
сопровождаться наи-
большим
эффектом оплодотворения в случае
одновременного мас-
сового
появления яиц и сперматозоидов.
Условия
для развития молоди в любом участке
гидросферы не
постоянны
во времени, и отсюда возникает та или
иная периодич-
ность
размножения, связанная главным образом
с сезонными и су-
точными
изменениями в поступлении солнечной
энергии. Особенно
261
четки
ритмы размножения в водоемах умеренных
и высоких широт,
где
колебания притока солнечной энергии
выражены наиболее
резко.
Однако и в тропических водоемах часто
наблюдается перио-
дичность
размножения, как это, например, отмечено
для многих
брюхоногих
и двустворчатых моллюсков в Индийском
океане. Пре-
имущественное
размножение в летнее время обнаружено
у некото-
рых
глубоководных животных, например у
офиуры Орютьзшт
1итат,
обитающей на глубине 3800 м. В Атлантике
на глубине
около
3 км моллюски Ьес1е11а
и Уо1(Ие11а
выметывают яйца соответ-
ственно
только весной и осенью, офиура О.
1]ип§та1
— только зи-
мой.
Морская звезда ЫерагйЫа
Ье1сНеп
зимой размножается деле-
нием,
а с начала лета переходит к половому,
обеспечивающему
расселение
животных (ОИезеп, Ьисаз, 1982). Рачки
Саттагиз
1а-
сиз1г1з
в высоких широтах начинают размножаться
летом при
12-часовом
световом дне, увеличивая численность
даже в условиях
низких
температур, но весной или осенью при
коротком световом
дне
не размножаются даже при благоприятных
температурах
(МагсЪ,
1982).
Сезонность
размножения с образованием планктоноядных
личи-
нок
свойственна многим глубоководным
бентонтам, в частности
ряду
иглокожих. Появление таких личинок,
совпадающее с макси-
мальным
развитием фитопланктона у поверхности,
свидетельствует
о
довольно быстром переносе органических
частиц в глубину, где
они
с ассоциированными с ними бактериями
используются личин-
ками
глубоководных животных. Приуроченность
периодов размно-
жения
к тем или иным сезонам определяется
главным образом
динамикой
температур. Через реакцию на этот
фактор у гидробион-
тов
в процессе эволюции выработались
наиболее рациональные
сроки
размножения применительно к геофизическим
циклам Земли.
Согласно
правилу известного океанолога Д.
Ортмана, большин-
ство
морских животных начинает размножаться
с наступлением
определенных
температур воды, причем для каждого
вида эта тем-
пература
специфична и константна. Некоторые
виды начинают
размножаться
не ‘При
определенной
температуре, а в случае резких
изменений
в ее ходе. Гидробионты, обитающие в
одном водоеме,
но
имеющие разное зоогеографическое
происхождение, обычно раз-
множаются
в разные сроки: холодноводные виды в
теплых водах —
с
наступлением температурного минимума,
а телловодныё виды в
холодных
водах—с наступлением максимума. У
видов космопо-
литов
период размножения обычно сильно
растянут.
Для
тепловодных форм, обитающих в холодных
морях, опти-
мальные
условия складываются в самое теплое
время, к которому
и
приурочено их размножение. Для
холодолюбивых форм, адапти-
рованных
к низким температурам, но обитающих в
теплых водах,
среда
становится оптимальной во время
наибольшего похолодания,
и
в эти сроки появляются молодые стадии.
Например, в Канда-
лакшском
заливе Белого моря арктические и
арктико-бореальные
формы
размножаются при более низких
температурах, чем бо-
реальные
и южно-бореальные виды, причем у каждой
из этих форм
262
диапазон
температур, с наступлением которых
начинается размно-
жение,
очень невелик, составляя около 1—2°С.
Космополитические
виды
значительную часть года размножаются
при температурах,
различающихся
на 16°С.
Сроки
размножения у видов с пелагическими
личинками корре-
лируют
с периодом массового появления
фитопланктона, которым
питается
молодь. Эта закономерность — одна из
самых универсаль-
ных,
поскольку пища представляет собой
ведущий фактор в обес-
печении
выживаемости личинок. Так, у морского
ежа Б^айета
8е1отиз,
обитающего
в тропической зоне и размножающегося
в те-
чение
всего года, гаметогенез в пределах
данной популяции проте-
кает
синхронно и наблюдается примерно
месячная периодичность
размножения;
по мере удаления от экватора сроки
размножения
ежа
все более ограничиваются и на широтах
выше 30° приходятся
только
на летние месяцы. Приуроченность
периодов размножения
к
тем или иным сезонам в наибольшей
степени обеспечивается сиг-
нальным
действием температур. Например, нерест
устриц Сга880-
81геа
уицШса
можно
задержать на любое время, если
температура
воды
достаточна для созревания гонад, но
ниже той, которая необ-
ходима
для вымета половых продутков.
Периодичность
размножения, связанная с чередованием
фаз лу-
ны,
бывает двух типов. При первом размножение
происходит не-
сколько
раз в течение лунного месяца, причем
наиболее интенсивно
в
новолуния или в полнолуния. При втором
типе периодичности
размножение
приурочено только к какой-либо одной
фазе луны,
т.
е. происходит один раз за лунный месяц.
Первый тип размноже-
ния
свойствен, например, полихете ЗрмогЫз
ЬогеаНв,
обитающей
у
берегов Франции. Ооциты этого червя
созревают 14 дней, столь-
ко
же времени продолжается инкубация
яиц. Каждую фазу луны
начинается
образование ооцитов, происходит
откладка яиц и вы-
мет
из них личинок. Подобный тип периодичности
наблюдается у
креветки
АпсЫзИоШез
апЩиеп815,
моллюска
Масота
ЬаШса
(рис.
44). В Кандалакшском заливе Белого моря
он свойствен
моллюскам
Ьасипа
й'маг1са1а, ЫНоппа ИИогеа,
МИИиз
ейиН8,
Масота
ЪаШса,
Муа агепаг'ш,
многим
усоногим, полихетам и игло-
кожим.
Периодичность
второго типа характерна для многих
полихет.
Например,
черви палоло Ейтсе
ь1гШз
на о-вах Самоа роятся
только
в 3-ю четверть Луны в октябре или ноябре
каждого года.
Отрывающиеся
от головных концов членики тела, набитые
поло-
выми
продуктами, начинают всплывать от дна
к поверхности в
О
ч 07 мин. В это время жители организуют
лов червей, употреб-
ляемых
в пищу. Роение полихеты Ос1оп1о$уШ8
епор1а
у
Бермудских
о-вов
происходит через три дня после новолуния
через 54 мин после
захода
солнца, повторяясь в это же время через
день или два.
А<еге18
V^^епз
в устье Темзы роится раз в году в первое
майское
новолуние.
Штормовая погода может задержать
роение этой поли-
хеты
на день или два, но не сдвигает его
относительно времени су-
ток.
263
Адгуст Сентябрь
Мни
лунного месяца
Рис.
44. Лунная периодичность в размножении.
А
— Масота
ЬаШса;
Б
— АпсЫзИгяёез
апИдиепз^з
(по Мооге, 1958):
/
— зрелые, 2
— отнерестившиеся особи
Природа
лунной периодичности размножения
неодинакова, но,
по-видимому,
в наибольшей степени связана с
чередованием перио-
дов
различной освещенности и сменой
приливов и отливов. Так,
б условиях
постоянного искусственного освещения
время роения
полихеты
РШупегеьз
йитепШ
утрачивает связь с чередованием
фаз
луны, как это наблюдается в природных
условиях; соответст-
вующим
чередованием периодов освещения у
этих червей можно
вызвать
самые разные ритмы роения. Некоторые
авторы считают,
что
действие прилива на организм сказывается
через изменение дав-
ления
воды. Следует отметить, что лунная
периодичность размно-
жения
отмечена для ряда форм в безливных
морях, например у
полихет
РШупегеьз
йитепШ
и Ыеге1з
гопа1а
в Черном море.
Изменения
в интенсивности размножения, связанные
с чередо-
ванием
дня и ночи, особенно характерны для
водорослей. Напри-
мер,
у массовых видов динофлагеллят Черного
моря РегШтит
и
СегаИит
максимум митозов наблюдается перед
рассветом, у диато-
мовых
Ехт1ае11а
и Ргогосегйгит
— во второй половине темного и
около
середины светлого периодов (Столбова
и др., 1982). Для
большинства
черноморских диатомовых минимум
делений прихо-
дится
на время с 22 до 4 ч.
У
некоторых быстроделящихся форм
наблюдаются два пика де-
лений:
один в утренние часы, другой во
второй'половине дня. Пре-
имущественное
деление в дневное время характерно
для многих
динофлагеллят.
264
Утренние
и вечерние ро-
ения
с последующей отклад-
кой
яиц характерны для мно- ^
гих
водных насекомых. Рое- ^ в
ние
кумовых раков, когда ~
они
из грунта поднимаются -г*,
в
толщу воды, наблюдается ^ | 4
вскоре
после захода солнца
и
прекращается с его восхо-
дом.
Утром выметываются |§ О
яйца
у актинии НуйгасИта
®
ескта1а.
На
рис. 45 видно, |
как
четко выражена суточ- =»• 2
ная
периодичность в размно- 0
жении
веслоногого рачка 12 24 12
Са1апиз
/'штагсЫсив. Время
суток, ч
Иногда
суточная перио-
дичность
размножения свя- Рис- 45
Суточный ритм откладки яиц у
„ „ „„„„„„„„„ Са1апи8
пптагстсиз
(по Мооге, 1958). Кри-
зана
не с изменением осве- вые
— число
отложенных яиц, столбики —
щенности,
а с чередованием число
самок, откладывающих яйца: А
— ин-
приливоь
и отливов, как это, тервал
между наблюдениями 1 ч;
Б
—
3
ч
например,
наблюдается в Бе-
лом
море у моллюсков Ьасипа
йыаг1са1а, ЫИоппа ИНогеа, Масо-
та
ЪаШса, Муа агепагьа
и
у некоторых других животных.
Четкий
суточный
ритм нереста обнаружен у ряда черноморских
рыб с пе-
лагической
икрой. У эстуарного краба РНу1горапореи8
Наггьвви
93%
случаев выпуска личинок приходится
на время максимума при-
лива—
предпочтительно в темное время суток
(Сгошп, 1982).
Темп
образования новых генераций у
представителей разных
видов
очень варьирует, причем, как правило,
с увеличением их
размеров
замедляется. Например, у инфузорий
такая зависимость
выражается
прямой в системе логарифмических
координат, т. е.
имеет
степенной характер. У некоторых бактерий
промежуток меж-
ду
последовательными делениями составляет
всего несколько ми-
нут,
а у простейших — часы и даже сутки.
Примерно такой же темп
размножения
водорослей. У мелких многоклеточных
частота гене-
раций
не превышает нескольких суток, у крупных
выражается
многими
сутками и месяцами.
Плодовитость.
Под плодовитостью популяции понимается
коли-
чество
яиц или других зачатков, образуемых
особями в единицу
времени.
Плодовитость популяции определяется
числом особей,
участвующих
в размножении, частотой этого процесса
и индиви-
дуальной
плодовитостью — количеством зачатков,
образуемых от-
дельными
организмами. Под абсолютной
плодовитостью
понимают
общее
число икринок, яиц или других зачатков,
образуемых орга-
низмом
в течение одного периода размножения.
Относительная
плодовитость
представляет
собой отношение абсолютной плодови-
тости
особи к массе ее тела. С увеличением
размеров гидробионтов
265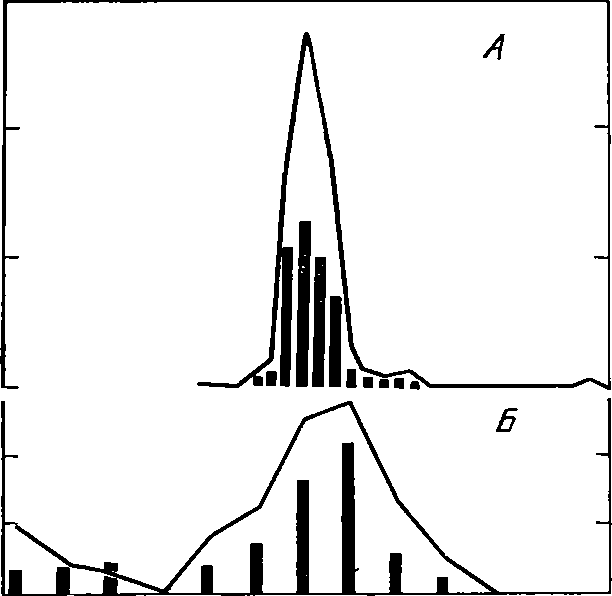
их
абсолютная плодовитость обычно
возрастает, а относительная
падает.
Формы
с длительным циклом развития, как
правило, более пло-
довиты,
чем формы с коротким циклом. Число
генераций, образуе-
мых
в течение жизни, и количество зачатков
за один срок размно-
жения,
с одной стороны, представляют собой
видовое свойство, с
другой
— зависят от внешних условий. По числу
образуемых гене-
раций
(поколений) различают формы, размножающиеся
в течение
жизни
один или несколько раз. Только один
раз в жизни размно-
жаются
и после этого гибнут рачки 8ер1еп1попаИз
и Рапаеиз
йюга-
гит,
поденки, веснянки, ручейники, многие
двукрылые, полихеты
Nеге^з
йшег8'1Со1ог
и N.
ь'1геп8,
ряд рыб, в частности большинство
дальневосточных
лососей, однолетние гидрофиты
(шильник,
наяда
малая, некоторые повойнички) и многие
другие гидроби-
онты.
Организмы,
размножающиеся несколько раз в жизни,
могут
быть
однолетними и многолетними. Среди
многолетних различают
формы,
дающие за один год одно, два или много
поколений. Число
генераций
за год у одного и того же вида зависит
от условий
существования,
в первую очередь температурных. Так,
амфиподы
АЧркагдиз
огстш Ю1ге1
и Оаттагиз
1асиз1г'18,
дающие в оз. Севан
4
поколения, в водоемах Прибайкалья и
Западной Сибири размно-
жаются
2 раза в год; рачок Апопух
пи@ах
в Баренцевом море раз-
множается
один раз, а в Белом море за 4—6 лет
образует кладки
не
более 4 раз. СЫгопотиз
рШтозиз
в арктических водоемах раз-
множается
один раз в 2 года, в Северной Европе
имеет в год одну
генерацию,
в озерах Подмосковья — 1—2, в прудах
Куйбышевской
области
— 2—3, а в водоемах дельты Волги — 4
генерации. С при-
ближением
к экватору все чаще встречаются
популяции рыб и дру-
гих
животных не с одноразовым, а с многократным
икрометанием
в
году, поскольку кормовая база для
молоди здесь более стабильна,
чем
в высоких широтах.
Чем
менее благоприятны условия для роста
и развития орга-
низмов,
тем позже наступает у них репродуктивная
зрелость и тем
реже
они образуют новые генерации. К факторам,
задерживающим
рост
и развитие организмов, помимо низких
температур, относятся
недостаток
пищи и кислорода, неблагоприятная
соленость и многие
другие.
Так, олигохета Иего
йщйа1а
не делилась в воде, содержащей
мг/л
02;
с повышением концентрации кислорода
до 4,6 и
мг/л
интервалы между делениями составляли
соответственно
15—16
дней и 3,2 дня. Сходные по размерам
моллюски ЫИоппа
оЫизШа
в Белом море обладают меньшей абсолютной
плодови-
тостью,
чем в Баренцевом, где кислородный
режим лучше. То же
самое
относится к рачкам Саттагиз
1аси81пз, С. тагтиз
и Не1ш-
гиз
ро1аг18.
В
очень большой степени плодовитость
зависит от размера об-
разуемых
зародышей. Чем они мельче, тем большее
их число мо-
жет
отрождать организм, но зато перспектива
достижения каждым
зародышем
репродуктивной зрелости сильно
сокращается. Кревет-
266
ка
Ра1аетопе1ез
Vа^^ап8
в пресных водах близ Неаполя отклады-
вает
20—25 яиц диаметром 1,3—1,4 мм, а в соленых
водах тех же
мест
образует яйца диаметром 0,7—0,8 мм, зато
их число состав-
ляет
100—450. У крупных особей плодовитость
обычно выше, чем
у
мелких. Например, у криля Еиркаиз1а
'зирегЬа
продукция яиц
прямо
пропорциональна длине самки и обратно
пропорциональна
стадии
ее зрелости (Оешз, ШЫпте, 1982).
Рачки
Роп(о^аттагиз
гоЪизШйез
размером 10—11 мм дают за
один
раз 24—27 яиц, а с увеличением длины
самок до 13—14 и
15—17
мм их плодовитость .возрастает
соответственно до 30—79 и
88—139
яиц (Касымов, 1960). У морских рачков
НаграсИсоШа
прослежена
прямая зависимость между объемом тела
и суммарным
объемом
откладываемых яиц. Для амфиподы
81е§осерка1из
т{1а(из
число
яиц (УУ) с увеличением длины самок (/)
выражается урав-
нением
N
=
0,000036 /4’03.
Плодовитость
организмов обычно тесно коррелирует
со сте-
пенью
заботы о потомстве. Как правило, у
червей, ракообразных,
моллюсков
и рыб, охраняющих свои яйца, зародышей
образуется
гораздо
меньше, чем у животных этих же групп,
не проявляющих
никакой
заботы об охране потомства. Так как у
водных животных
преобладает
наружное оплодотворение, то для них в
целом харак-
терна
относительно высокая плодовитость.
Шансы на встречу гамет
при
наружном оплодотворении сравнительно
невысоки, и гидро-
бионтам
приходится компенсировать этот
биологический минус по-
вышением
числа выметываемых гамет. Так, у морского
ежа 3(гоп-
цу1осеп1го1из
с1гоеЬасЫеп$1$,
обитающего в Белом море, на долю
гонад
в период половой зрелости приходится
около 15,8% от массы
всего
тела, а после вымета половых продуктов
— 3,8%. У морской
звезды
Аз1епаз
гиЬепз
масса гонад после икрометания снижается
с
22 до 0,8%, примерно такое же соотношение
характерно для го-
лотурии
Сиситапа
[гопёоза.
Моллюск 81оторпеизре8
ьаг1о1аг1з
за
период
икрометания теряет до 23% массы своего
тела, ЫИоппа
ЦИогеа
— 25—32, рачок Ва1апиз
Ъа1апоЫе8
— 30—35%.
Согласно
Г. В. Никольскому (1974), по мере роста рыб
до опре-
деленного
возраста плодовитость возрастает, а
затем снижается.
Сначала
уменьшается относительная плодовитость,
а затем иногда
и
абсолютная. Популяционная плодовитость
увеличивается также
в
результате более раннего созревания
особей, причем в популя-
циях
одного и того же вида и близких видов
формы высоких широт
обычно
менее плодовиты. В первую очередь это
связано с большим
прессом
хищников в низших широтах. Перечисленные
особенности
характерны
для очень многих гидробионтов.
Смертность
и выживаемость
Численность*
особей каждой генерации непрерывно
снижается
вследствие
естественного отмирания, в результате
уничтожения
другими
организмами и гибели из-за неблагоприятных
контактов
267
с
абиотическим окружением. Темп этого
снижения характеризует
смертность
популяции. Величина, обратная смертности,
называется
выживаемостью
и характеризуется числом особей,
доживающих до
того
или иного возраста. Адаптации к снижению
смертности (по-
вышение
выживаемости) можно подразделить на
те, которые сни-
жают
гибель зародышей, и те, которые направлены
на сохранение
особей
в постэмбриональный период.
Смертность.
Различают смертность за тот или иной
срок (АЛ^),
за
единицу времени (ДЛ7Д0 и уровень
смертности (АN^А^N).
Показателем
смертности в известной мере может
служить плодо-
витость
популяций. Если средняя численность
последних на протя-
жении
значительных отрезков времени остается
сходной, это зна-
чит,
что отмирает столько организмов,
сколько их образуется.
Плодовитость
можно рассматривать как адаптацию к
той или иной
смертности.
У некоторых гидробионтов она выражается
астрономи-
ческими
цифрами: луна-рыба, например, за один
раз выметывает
до
300 млн. икринок. Огромнейших величин
достигает рождаемость
в
популяциях мелких, партеногенетически
размножающихся рако-
образных,
когда каждая особь образует яйца с.
интервалом в 2—3
дня.
Еще выше темп размножения бактерий и
одноклеточных водо-
рослей
и соответственно выше их смертность.
В популяциях мелких
гидробионтов
смертность в основном определяется
выедаемостыо
особей
и их гибелью в результате неблагоприятных
условий абиоти-
ческой
среды.
В
популяциях крупных животных ведущую
роль может приобре-
тать
отмирание старых особей.
Величина
смертности не характеризует специфику
гибели орга-
низмов
разных возрастных групп в популяции.
Эту специфику от-
ражает
кривая
смертности,
показывающая степень снижения
чис-
ленности
особей данной генерации с момента
отрождения до конца
жизненного
цикла. Каждая популяция имеет типичную
для нее
кривую
смертности, которая несет отпечаток
многих сторон эко-
логии
вида (рис. 46).
Для
популяций гидробионтов наиболее обычен
тип смертности,
характеризующийся
снижением гибели организмов по мере
их роста
и
усиления эффективности различных
защитных средств. Часто
такой
характер смертности хорошо описывается
уравнением Л^ =
=Л/0е-**,
где N0
и N1
— соответственно численность пополнения
в
начальный
момент и в возрасте I,
к
— константа, называемая коэф-
фициентом
элиминации, е
— основание натуральных логарифмов.
Например,
по данным М. М. Камшилова (1955), в
Баренцевом
море
смертность рачка Са1апив
/ШтагсЫсиз
в 1950—1954 гг. хоро-
шо
выражалась уравнением Л^ = 695-Ю-0-00814
* (возраст в сутках,
численность
— в экз./м3).
Та
или иная смертность на разных стадиях
развития связана
не
только с возрастными изменениями
конституциональной защи-
щенности
организмов (увеличение размеров,
скорости движения,
образование
защитных структур и т. д.). Существенное
значение
может
иметь смена образа жизни на той или
иной стадии развития,
268
например
переход личинок
из
толщи воды на грунт
или
заглубление в него. По-
вышенная
смертность часто
обусловлена
не снижением
защитных
свойств организ-
ма,
а увеличением числа его
потребителей
или резким
ухудшением
абиотической
среды.
Так, смертность в по-
пуляциях
гребневика боли-
нопсиса
резко повышается с
массовым
появлением в во-
де
его потребителя — греб-
невика
берое, численность
популяций
эвфаузиид силь-
но
снижается, когда в райо-
не
их нахождения появляют-
ся
киты. Частичная или пол-
ная
гибель популяций может
наблюдаться
после промер-
зания,
высыхания водоемов,
резкого
ухудшения условий дыхания (заморы),
при сильном изме-
нении
солености среды и некоторых других
катастрофических ситу-
ациях.
Выживаемость
зародышей. Снижение смертности
зародышей
обеспечивается
главным образом благоприятными
респираторными
температурными
условиями, а также защитой от уничтожения
дру-
гими
организмами. Благоприятные условия
для дыхания зароды-
шей
обычно создаются в местах, богатых
кислородом, проточ-
ностью
находящейся вокруг них воды. Улучшению
газообмена спо-
собствуют
малые размеры яиц (увеличение
относительной поверх-
ности),
а также образование различных дыхательных
пигментов.
Универсальный
принцип улучшения условий дыхания
пелагических
яиц
—:
размещение их в более аэрированных
поверхностных слоях
воды.
В одних случаях это достигается
снижением плотности яиц,
в
других — прикреплением их вблизи
поверхности воды. В верхних
горизонтах
воды плавают яйца многих полихет,
иглокожих, насека-
мых,
икра многих кефалиевых, скумбриевых,
ставридовых и ряда
других
рыб.
Если
яйца тяжелее воды, то они удерживаются
в поверхностном
слое,
прикрепляясь к тем или иным предметам.
Гидробионты, от-
кладывающие
яйца на дне, выбирают участки с наилучшим
кис-
лородным
режимом. Ныряя в воду, поденки ВаеНз
приклеивают
яйца
к камням на заметном течении, в таких
же местах встречают-
ся
кладки многих брюхоногих моллюсков и
некоторых рыб, в ча-
стности
осетровых. К песчинкам на течении
прикрепляются коконы
многих
олигохет. Некоторые насекомые откладывают
яйца внутрь
живых
растений, в . воздухоносных тканях
которых создаются
Продолжительность
жизни. В
%
от
'длительности
Всей
жизни
Рис.
46. Кривые смертности некоторых
гид-
робионтов:
/
— устрицы, 2
— гидра, 3
— водные млекопитаю-
щие
269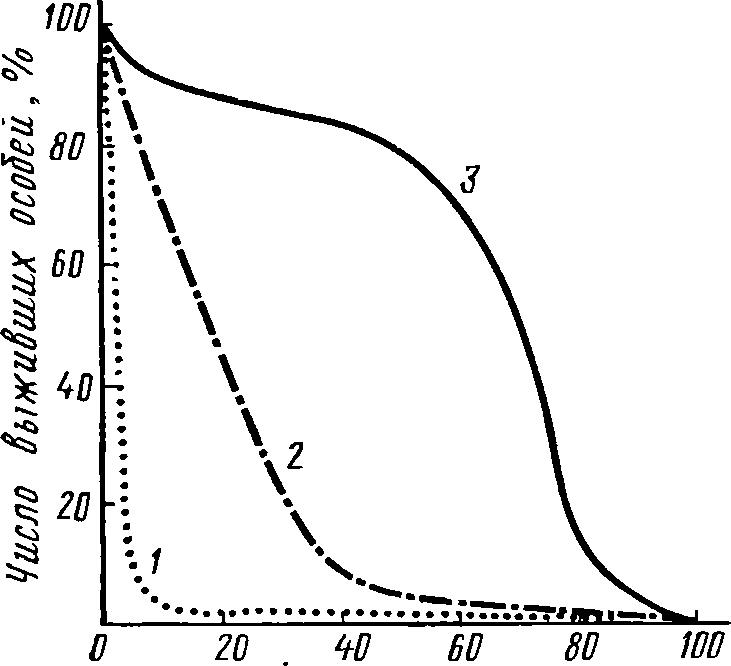
идеальные
условия дыхания. Например, самки
стрекозы Аезскпа
и
Ье81в8
погружаются в воду и, оставаясь здесь
до 25 мин, с по-
мощью
яйцеклада втыкают яйца в воздухоносную
ткань растений..
Погружают
в растения свои яйца вилохвостка
ЗтМкигШез,
жук
ОуИзсиз
таг^таИз,
клоп ЫоЬопесЫ
&1аиса
и многие другие насе-
комые.
В
ряде случаев условия газообмена
зародышей улучшаются за
счет
прикрепления их к родительскому или
иному организму, обла-
дающему
активным движением. Благодаря
перемещениям живот-
ных
яйца постоянно омываются током свежей
воды, что улучшает
их
снабжение кислородом и способствует
удалению выделяемого
С02.
В частности, омываются во время _
движения родительских
организмов
яйца низших и высших ракообразных,
прикрепляемые
к
ножкам, к концу брюшка или иным частям
тела. Если яйца ра-
кообразных
развиваются в выводковых камерах,
последние могут
попеременно
сокращаться и увеличиваться в объеме
и таким об-
разом
вода периодически обновляется. У рыбки
Ародоп
икра вы-
нашивается
в открытой ротовой полости самца,
сквозь которую все
время
проходит ток воды. У рыб Р1а1уз1аски8
со1у1еркогиз
икра
находится
в специальной выводковой камере на
брюхе, а у Таску-
8иги8
ЬагЬи.8
— даже в желудке. Рыбка СогергосЫз
прикрепляет
икру
под панцирь камчатского краба, и он,
передвигаясь, освежа-
ет
воду вокруг зародышей.
Иногда
смена воды вокруг развивающихся яиц
наблюдается *г
у
малоподвижных животных. У моллюсков
Апойоп^а,
Ито
и ряда
других
яйца развиваются в жабрах, сквозь
которые непрерывно'
проходит
ток воды. Самки червей Ыегеьз,
Нурата ьпьаИйа
откла-
дывают
яйца в своих норках и колебаниями тела
поддерживают в
них
проточность; в трубках с проточной
водой откладывают яйца
и
погонофоры. Самки осьминога Ос1орив
уи1&апз
периодически
освежают
воду вокруг яиц, направляя на них струю
из сифона; во
избежание
загрязнения воды и снижения в ней
концентрации кис-
лорода
самки почти совсем не питаются во время
«высиживания»
и
к концу развития яиц часто погибают.
Самцы колюшки время от
времени
нагнетают в гнездо с икрой свежую воду
движением груд-
ных
плавников.
Эмбриональное
развитие ускоряется с повышением
температу-
ры.
Поэтому многие гидробионты совершают
миграции для отклад-
ки
яиц в местах с благоприятным термическим
режимом. Очень
часто
яйца инкубируются в верхних, более
прогреваемых слоях,
воды.
Сокращение длительности эмбрионального
развития полезно
для
вида не только с точки зрения повышения
темпа воспроизвод-
ства,
но и потому, что в зародышевом состоянии
организмы осо-
бенно
уязвимы.
Сохранность
яиц в огромной степени определяется
их защищен-
ностью
от врагов, которая достигается качествами
икры, затруд-
няющими
ее поедание, укрытием от врагов или
активной охраной
родителями.
Ограничивает обнаружение яиц высокая
прозрачность,
свойственная
кладкам моллюсков и насекомых, икринкам
рыб и.
270
яйцам
ракообразных. Как правило, яйца
гидробионтов одеты сли-
зистой
оболочкой или заключены в кладку,
окруженную слизью.
С
одной стороны, слизь может обладать
бактерицидными свойст-
вами
и защищать яйца от микроорганизмов, с
другой — затрудняет
схватывание
и поедание их животными.
Многие
гидробионты, помещая яйца в укрытия,
снижают или
полностью
исключают их поедание. Под защитой
родительских ор-
ганизмов
находятся яйца ракообразных, прикрепленные
к ножкам
или
другим частям тела животных, яйца
моллюсков, развивающие-
ся
в их жабрах, яйца полихет и погонофор,
откладываемые в труб-
ках.
Многочисленны примеры активной охраны
яиц животными.
Например,
сидят возле них, отгоняя морских звезд
и других вра-
гов,
самки осьминогов; находится около яиц
в воздушном колоколе
самка
водяного паука Аг^угопе1а
ациаИса.
Активно охраняют икру,
отгоняя
врагов, многие рыбы.
Голожаберные
моллюски Р1егосо1ща
1апИпа
группами окружа-
ют
яйцекладки, закрывая их своими телами;
наличие нематоцистов
усиливает
защиту от врагов, а присутствие
зооксантелл позволяет
реже
удаляться для поиска пищи (Козе, 1982).
Закрывают своими
телами
отложенные яйца и некоторые морские
звезды, в частности
Ьер1а.$1ег1аз.
Наибольшая
выживаемость зародышей наблюдается
в случае
живорождения.
Чаще оно встречается у организмов,
обитающих в
крайних
условиях абиотической среды, в частности
в 'водоемах с
низкими
температурами. Например, из 57 живородящих
офиур
только
7 известны для тропических мелководий
(НепсНег, 1979).
В
сходном плане следует оценивать смену
типов вылупления мо-
лоди.
Так, с продвижением в высокие широты
у моллюсков Мипсь
<1ае
вылупление на стадии пелагической
личинки — велигера заме-
щается
образованием ювенильных донных особей.
Виды
актиний семейства ТаеНйае, живущие в
арктических усло-
виях,
имеют специальные выводковые камеры,
отсутствующие у
представителей
этой группы, населяющих более теплые
водоемы;
актиния
ТаеНпа
сгаззкогтз
у берегов Франции яйцекладущая, а
у
о. Шпицберген — живородящая. Рачок
Аг1ет1а
заИпа
в условиях,
препятствующих
образованию гемоглобина, и при недостатке
в
пище
хлорофилла становится живородящим,
хотя в обычных усло-
виях
откладывает яйца.
Часто
развитие яиц задерживается, если
условия в водоеме
неблагоприятны
для вылупляющейся молоди. Таково
значение по-
коящихся
яиц многих листоногих и веслоногих
рачков, ресничных
червей,
коловраток, нематод. Покоящиеся яйца
обладают очень вы-
сокой
устойчивостью к действию крайних
температур, высыханию
и
другим неблагоприятным факторам.
Например, яйца щитня
Ариз
в сухом грунте могут выдерживать
повышение температуры
до
42 и даже до 80°С, а зимой хорошо выносят
промерзание.
Крайне
устойчивы к высыханию, прогреванию и
промерзанию зим-
ние
яйца ресничных червей Мезоз1ота
и Во1Нотезоз1ота,
коловра-
ток
Ро1уаг1кга
рШур1ега,
жабонога ерШигиз
и многих других жи-
271
Рис.
47. Относительное число видов
переднежаберных (1)
и пластинчатожабер-
ных
(2)
моллюсков, имеющих пелагических
личинок, в процентах от общего чис-
ла
видов на разных широтах
вотных.
Высокой устойчивостью к внешним
воздействиям отлича-
ются
споры и семена большинства водных
растений.
Выживаемость
в постэмбриональный период.
Выживаемость
особей
в этот период прежде всего достигается
маневренным
использованием
разных способов развития. Как правило,
в благо-
приятных
условиях чаще встречаются виды с
непрямым развитием,
а
в более суровых —с прямым. Развитие с
метаморфозом, т. е. су-
ществование
вида в форме особей, резко отличных
друг от друга
по
своему строению и потребностям, имеет
большое значение для
увеличения
численности популяций. Например,
наличие планкто-
трофных
пелагических личинок у бентосных
животных позволяет
им
использовать корм, не потребляемый
взрослыми организмами,
а
ранний переход личинок к самостоятельному
питанию обусловлен
малым
количеством желтка в яйцах, что в свою
очередь создает
предпосылку
для повышения плодовитости особей. С
другой сто-
роны,
молодь, развивающаяся с метаморфозом,
менее сформирова-
на,
и выживаемость неокрепших личинок
низкая. Пелагические ли-
чинки
бентосных животных в конце развития
оседают на дно, кото-
рое
в данном месте далеко не всегда
благоприятно для жизни, и
огромное
количество молоди гибнет. Таким образом,
обе формы
развития
выгодны и уязвимы, вследствие чего в
разной степени
свойственны
видам, обитающим в неодинаковых
условиях.
Согласно
правилу Г. Торсона, с продвижением от
экватора к
полюсам
и от прибрежных мелководий к батиальным
глубинам не-
272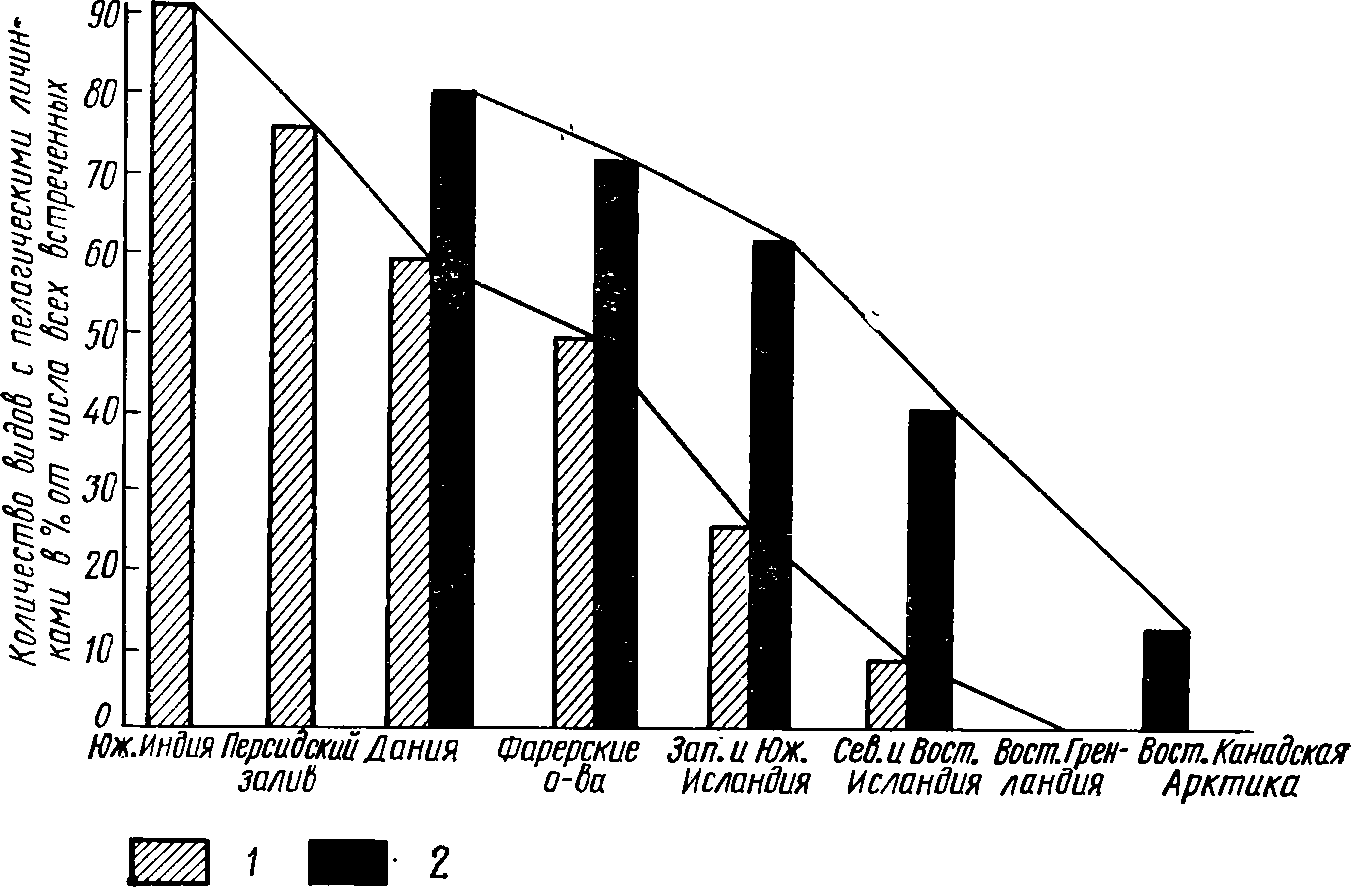
прямое
развитие, с образованием пелагических
личинок, наблю-
дается
все реже. Наиболее четко эта закономерность
прослежена
на
переднежаберных моллюсках, у которых
относительное количе-
ство
видов с пелагическими личинками с
(продвижением от тропи-
ков
к Арктике снижается с 91% До
0
(рис. 47). Почти столь же
четко
рассматриваемая закономерность
выражена у двустворчатых
моллюсков.
Число их видов с пелагическими личинками
снижается
с
продвижением от побережья Дании к
Арктике с 79 до 13. В целом
е
прибрежье тропических и субтропических
морей пелагические
личинки
свойственны представителям 90—95% всех
видов бентос-
ных
животных, в водах бореальных — 65—70, а
в полярных — око-
ло
5% (Регек, 1961). Из этого общего числа
видов примерно 10%
приходится
на долю форм с лецитотрофными
личинками, живущи-
ми
за счет желтка и не потребляющими
планктон, и около 90% —
на
долю форм с личинками планктотрофными.
Лецитотрофные ли-
чинки,
имеющиеся, например, у губок 8усоп
1епсозо1етз,
моллюска
Ыиси1а
ргохьта
и ряда других животных, пребывают в
планктоне
в
течение нескольких часов или дней.
Планктотрофные личинки
находятся
в толще воды от нескольких недель до
2—3 месяцев и
более.
Важный
фактор, определяющий распространение
бентосных
форм
с пелагическими личинками, — степень
обеспеченности их
пищей.
В тропиках количество пищи (фитопланктона)
для планк-
тотрофных
пелагических личинок на протяжении
всего года до-
вольно
одинаково. С продвижением к полюсам
период развития
фитопланктона
укорачивается, и приурочить выход
пелагических
личинок
ко времени, когда для них есть пища,
биологически слож-
нее.
Поэтому экологическая выгода непрямого
развития с продви-
жением
к полюсам снижается, и виды, которым
оно свойственно,
становятся
все более редкими. Немаловажное значение
имеет и
суровость
температурных условий, возрастающая
с продвижением
в
высокие широты.
Зависимость
способа развития от экологических-
условий ка-
сается
не только крупных систематических
единиц, но и видов од-
ного
рода. Например, сублиторальный моллюск
ЫМоппа
ИИогеа
развивается
с пелагической личинкой, литоральная
гшИз
имеет
прямое
развитие, а верхнелиторальная пепШйез
— живородя-
щая.
Большинство видов имеет один тип
развития 'на всем протя-
жении
своего ареала, однако у некоторых
гидробионтов, например
у
многих полихет и моллюсков с ареалом,
захватывающим резко
различные
районы моря, отдельные особи вида,
живущие в неоди-
наковых
экологических условиях, могут обладать
разными форма-
ми
развития. Варьирование прямого и
непрямого развития наблю-
дается
у брюхоногих, полихет Реппеге1з
сиИг^ега
и Ыегеьз
соз1аеа,
некоторых
моллюсков-древоточцев.
Большое
значение для выживания особей имеет
регулирование
темпа
развития. Например, при отсутствии
подходящего субстрата
метаморфоз
личинок донных животных может
задерживаться на
несколько
дней или даже недель. Это имеет огромное
значение, так
273
как
увеличивается возможность выбора
подходящего субстрата и
соответственно
повышается выживаемость личинок.
Когда
среда особенно неблагоприятна, многие
гидробионты пре-
кращают
активное существование, приобретают
в это время значи-
тельно
большую устойчивость к внешним
воздействиям и за счет
снижения
темпа развития повышают свою выживаемость.
Выживаемость
особей в сильнейшей степени зависит
от харак-
тера
и темпа их роста. В соответствии с
конкретными условиями
среды
и состоянием популяции скорость роста
и его характер у осо-
бей
могут существенно меняться, обеспечивая
маневренность
использования
жизненных ресурсов и наибольшую
выживаемость
организмов.
Последняя, как правило, повышается с
ускорением
роста
и более ранним достижением крупных
размеров.
Когда
условия благоприятны, темп роста
повышается, и попу-
ляция,
используя сложившуюся обстановку,
быстро увеличивает
свою
биомассу. В неблагоприятных условиях
рост особей замед-
ляется
и популяция может полностью выключаться
из процесса
своего
воспроизводства. Так возникает
периодичность роста у осо-
бей
популяций, обитающих не только в
высоких, но и в низких ши-
ротах.
Например, у тропических кораллов рост
замедляется в пе-
риод
выпадения дождей, когда вода более
мутная, и во время
ослабления
пассатных ветров, влекущего за собой
снижение про-
точности
воды с соответствующим ухудшением
условий дыхания и
питания.
С продвижением к полюсам и нарастанием
сезонных
контрастов
в природе периодичность роста в
популяциях гидроби-
онтов
приобретает все более массовый и резяе
выраженный харак-
тер,
особенно в континентальных водоемах,
где условия среды ме-
няются
особенно сильно.
Рост
популяций
Под
ростом популяций понимают увеличение
их численности
(или
биомассы) во времени: ЛУ=ЛУ(/). Если
смертность преобла-
дает
над рождаемостью, рост становится
отрицательным. Разница
между
величиной популяций в начале и конце
какого-то промежут-
ка
времени (АЩ
называется приростом,
который может быть как
положительным,
так и отрицательным (убыль).
Прирост за едини-
цу
времени (ДЛ^/Д/) характеризует скорость
роста
популяций. Она
может
быть постоянной (неограниченный рост)
или затухающей,
когда
с увеличением численности особей
условия их существова-
ния
ухудшаются и темп воспроизводства
падает. Иногда скорость
роста
сначала возрастает с увеличением
плотности популяций, а
потом
снижается (тип Алли). Первый случай в
естественных по-
пуляциях
гидробионтов наблюдается редко, второй
характерен для
роста
популяций, не достигших стационарного
состояния. После его
достижения
происходит осцилляция
(колебание) численности отно-
сительно
некоторой величины, которая, однако,
может существенно
варьировать
в связи с различными изменениями в
водоемах.
274
Неограниченный
рост. Если пищевые, пространственные
и дру-
гие
жизненные ресурсы популяции безграничны,
а смертность с воз-
растанием
численности особей не увеличивается,
скорость роста
популяции
описывается уравнением
где
г
— мгновенная скорость роста популяции.
Кривая роста в этом
случае
имеет Л-образный характер и описывается
уравнением
Ы1=Ы0ег{,
где Ы0
и Ыг
— численность особей начальная и в
момент /.
Коэффициент
г
— характерная для вида величина,
отражающая
разность
между рождаемостью и смертностью.
Уравнение
(1) отражает потенциальные возможности
увеличе-
ния
численности популяций. В этом плане
коэффициент г
(«специ-
фическая
скорость естественного увеличения»)
имеет определенную
сравнительную
ценность. В какой-то степени уравнение
(1) при-
годно
для описания роста популяций (например,
бактерии) в лабо-
раторных
условиях, когда факторы, тормозящие
размножение,
устраняются
в эксперименте (пересевы и др.). В
водоемах, где
запасы
пищи ограничены, а также лимитированы
поступление кис-
лорода
и скорость других процессов, обеспечивающих
существова-
ние
популяции, экспоненциальный
(неограниченный) рост в тече-
ние
сколько-нибудь длительного времени
невозможен. К этому
следует
добавить, что соотношение рождаемости
и смертности,
определяющее
коэффициент г,
сильно изменяется даже в постоян-
ных
условиях среды (например, плодовитость
особей связана с их
возрастом).
Таким образом, экспоненциальный рост
— абстракция,
помогающая
моделировать изменение численности
популяций в не-
которых
условиях. В частности, знание коэффициента
г
очень важ-
но
для прогноза возможных вспышек
численности и определения
потенциальной
способности популяции к росту.
Затухающий
рост. В тех случаях, когда с увеличением
популя-
ции
дальнейшие возможности ее роста
становятся все более огра-
ниченными,
изменение численности описывается
«логистической»
кривой,
имеющей 5-образную форму. Она описывает
рост популя-
ций
животных, располагающих ограниченными,
но восполняемыми
жизненными
ресурсами (пища и др.). Вполне понятно,
что при не-
восполнимости
запасов пищи, кислорода и других средств
жизни
численность
популяции через какое-то время должна
понизиться
до
0. Логистическая кривая, предложенная
Ферхюльстом (1845) и
Перлем
(1952), описывается уравнением
где
к
— возможная максимальная численность
популяции. Это
уравнение
(2) похоже на первое (1), но в него включен
множитель
(к—Л/)/&,
выражающий «сопротивление среды»
приросту. Ассимп-
тота
численности, к которой приближается
кривая, соответствует
пределу
численности популяции, возможной в
данных условиях.
и.14 л
т
=
г/у, откуда г
1п
N2
— 1п
(1)
275![]()
Обычно
численность сначала нарастает медленно
(лаг-фаза),
затем
ускоряется (лог-фаза), снова замедляется
(фаза отрицатель-
ного
ускорения) и, наконец, наступает
состояние, когда прибыль и
убыль
особей приблизительно уравновешивают
друг друга (фаза
равновесия).
Рост
численности популяции наблюдается,
когда к исходному
моменту
она имеет меньше особей по сравнению
с их количеством,
возможным
в данных условиях. Такая ситуация
складывается, на-
пример,
ранней весной во многих водоемах с
почти отсутствующим
фитопланктоном,
который затем 'начинает бурно развиваться,
да-
вая
весеннюю вспышку. Когда величина
популяции достигает изве-
стного
уровня, рост ее прекращается, заменяясь
колебаниями
(осцилляцией)
в обе стороны от некоторой средней
величины, если
не
произойдет каких-либо изменений в
среде, в соответствии с чем
установится
новый уровень численности и биомассы
особей.
Рост
популяции, выражаемый в показателях
численности и био-
массы,
нередко носит разнонаправленный
характер. Например,
численность
байкальских рачков Мгстгориз
роззо1ски
и СтеНпог-
<1ез
}азааШз
с мая по сентябрь неуклонно снижается,
а средняя
масса
возрастает, вследствие чего биомасса
популяции в летние
месяцы
не только не падает, но даже несколько
увеличивается.
Динамика
численности и биомассы
популяций
Основная
закономерность самовоспроизводства
популяций за-
ключается
в постоянной тенденции к повышению
численности и
биомассы
особей. Их среднее количество может
быть высоким при
небольших
или значительных колебаниях уровня
численности по-
пуляций,
вплоть до таких, когда вспышки массового
развития гид-
робионтов
сменяются периодами их существования
только в форме
покоящихся
зачатков. Среда, меняясь, становится
то более, то ме-
нее
благоприятной для данной популяции,
и ее численность при-
способительно
меняется, чем достигается максимальное
использо-
вание
благоприятных условий, складывающихся
в те или иные
отрезки
времени в отдельных участках ареала.
Параллельно уве-
личению
численности особей в благоприятных
условиях ускоряется
их
рост, и в результате суммарная биомасса
популяции резко воз-
растает.
С ухудшением условий существования
численность и био-
масса
популяций падают, т. е. их воспроизводство
сужается до
степени,
соответствующей конкретным возможностям.
Таким обра-
зом,
динамика популяций видов имеет
приспособительное значение,
и
ее характер интегрально отражает
специфику взаимосвязи попу-
ляции
со средой. Динамика популяций есть как
бы фокус, в кото-
ром
концентрируются все ее экологические
особенности.
Тип
динамики популяций определяется
характером взаимодей-
ствия
зависимых друг от друга процессов
размножения, роста,
развития
и убыли особей. Так как убыль и пополнение
популяций
в
разные отрезки времени имеют разную
количественную выражен-
276
ность,
численность вида обнаруживает те или
иные колебания, или
флуктуацию.
Флуктуации тем сильнее, чем короче
жизненный цикл
организмов
и чем менее стабильна их среда. С
увеличением дли-
тельности
жизни гидробионтов их популяции
образуются все боль-
шим
числом разновозрастных поколений, и
резкое сокращение чис-
ленности
одного из них нивелируется урожайностью
других. У форм
с
коротким циклом неурожайность одного
поколения приводит к
резкому
сокращению численности популяций,
поскольку не компен-
сируется
особями других поколений.
Основной
фактор, ограничивающий нарастание
популяций, —
дефицит
пищи, что не умаляет роли других
факторов: недостатка
кислорода,
суровых температурных условий, выедания,
заражен-
ности
паразитами и многих других. Но количество
пищи — наибо-
лее
универсальный регулятор численности,
так как организмы мо-
гут
приспособиться ко многим неблагоприятным
условиям, но
только
не к отсутствию пищи. Поэтому на
ухудшение кормовых
условий
популяция реагирует снижением
численности. Например,
с
исчерпанием биогенов, нужных для
питания водорослей, числен-
ность
их после весенней вспышки начинает
резко падать, а вслед
за
этим уменьшаются популяции рачков и
других организмов, жи-
вущих
за счет фитопланктона.
Для
рыб Г. В. Никольский (1982) отмечает
следующие основ-
ные
пути приспособительного увеличения
популяций с улучшением
условий
питания: увеличение скорости роста,
более ранняя поло-
возрелость,
увеличение плодовитости одноразмерных
особей; повы-
шение
жирности, снижение смертности более
упитанных особей;
снижение
поедания собственной молоди; уменьшение
смертности
личинок
на первых этапах активного питания
(икра содержит боль-
ше
жира, и молодь переходит на внешнее
питание более окрепшей).
В
случае ухудшения условий питания
наблюдается обратная кар-
тина.
Отмеченные закономерности характерны
и для многих бес-
позвоночных.
К, важнейшим факторам, определяющим
величину
популяции,
помимо пищи относятся выедаемость
особей, особенно
в
случае их слабой защищенности от
врагов, а также конкретные
абиотические
условия, складывающиеся в водоемах.
Численность
и биомасса популяций могут меняться
периодически
или
непериодически. Второй тип изменений
связан со сдвигами в
условиях
существования, не повторяющимися во
времени с опре-
деленной
последовательностью. Резкие перемены
освещенности и
температуры
в смежные дни, выпадение осадков, штормы
и другие
события,
не связанные определенными сроками,
могут содейство-
вать
воспроизводству популяции или
обусловливать ее убыль.
Сходное
значение имеет изменение режима
водоемов под влиянием
человека:
зарегулирование стока рек, изъятие
воды на орошение,
подогрев
вод тепловыми станциями, загрязнение
промышленными
стоками
и др.
Более
существенны изменения численности и
биомассы популя-
ций,
имеющие колебательный характер. К ним
прежде всего отно-
сятся
те, которые связаны с геофизическими
циклами (суточная,
277
сезонная
и годовая динамика), циклами Луны и
Солнца. Очень
часто
они возникают при взаимодействии
зависимых друг от друга
популяций
(хищник — жертва, хозяин — паразит и
др.) или перио-
дических
воздействий человека.
Суточная
динамика. Суточные изменения численности
и биомас-
сы
прослеживаются в популяциях мелких
организмов с очень крат-
ким
жизненным циклом — бактерий, одноклеточных
водорослей и
простейших,
у которых имеется суточный ритм
размножения или
смертности.
Так, наибольшее количество водорослей
в Черном море
наблюдается
в 21 ч, затем их становится меньше, и
новое увели-
чение
численности начинается в 5 ч. Уменьшение
численности по-
пуляций
водорослей ночью связано с энергичным
выеданием их
рачками,
поднимающимися для питания из глубин
к поверхности
в
темное время суток. Нарастание количества
водорослей в днев-
ное
время обусловливается размножением,
превышающим убыль.
Для
большинства видов максимум делящихся
клеток приходится на
утренние
часы, днем интенсивность размножения
у разных видов
колеблется,
но у всех форм она минимальна в ночное
время — от
22
до 4 ч.
В
результате размножения преимущественно
в светлое время
суток
и гибели в темное суммарное количество
водорослей днем
часто
в 2—3 раза больше, чем ночью. Так, ночью
в Черном море в
2
раза снижается численность диатомовой
Ехт'шеИа
согопа1а
и
других представителей этого рода.
Численность популяции 01-
поНа2е11а1а
и ЗШсоНа^еГЫа в дневное время в 3—4 раза
больше,
чем
ночью.
Суточная
динамика численности хорошо прослеживается
и у
бактерий.
Например, в районах апвеллингов
Атлантики концентра-
ция
бактерий в слое 0—100 м днем достигала
1 млн./мл, а ночью —
0,7
млн./мл. Вне зон апвеллингов численность
бактерий равнялась
соответственно
138 и 75 тыс./мл; снижение количества
микроорга-
низмов
ночью объясняется их выеданием
мигрирующим зоопланкто-
ном
(Лебедева, 1976). Известное значение имеет
и ингибирование
размножения
бактерий метаболитами активно делящихся
днем во-
дорослей.
Суточным ритмом питания ряда животных
может обу-
словливаться
соответствующая динамика численности
простей-
ших
и других мелких гидробионтов с коротким
циклом разви-
тия.
Сезонная
динамика. Сезонные колебания численности
и биомас-
сы
популяций водных организмов в основном
связаны с изменением
интенсивности
солнечной радиации как непосредственного
источни-
ка
энергии для фотосинтезирующих растений
и как фактора, влияю-
щего
на термику водоемов. Изменения в
количестве падающего
света,
обусловливая периодичность развития
водорослей, опреде-
ляют
и цикличность динамики численности
животных, питающихся
растениями.
Сезонная
динамика планктонных водорослей.
Динамика
численности и биомассы водорослей
определяется глав-
ным
образом сезонными изменениями
освещенности, количества
278
питательных
солей, температуры, вертикальной
устойчивости вод-
ных
масс, а также ритмом выедания водорослей
животными. Се-
зонные
изменения освещенности увеличиваются
в направлении
высоких
широт, и соответственно этому резче
разница в количест-
венном
богатстве фитопланктона с переходом
от весны к лету, осени
и
зиме. Так, на протяжении года численность
диатомовых водорос-
лей
у юго-западных берегов Англии изменяется
приблизитель-
но
в 500 раз, а у северных берегов Австралии
— примерно
в
10 раз.
По
мере развития фитопланктона вода
обедняется питательны-
ми
солями, и численность водорослей
начинает снижаться. После-
дующее
накопление биогенов сопровождается
новым увеличением
количества
водорослей. В тех случаях, когда
количество питатель-
ных
солей имеет не один, а несколько
максимумов на протяжении
периода
вегетации, в развитии фитопланктона
наблюдается соот-
ветствующее
число вспышек. Как правило, в тропических
и умерен-
ных
морях прослеживаются два пика численности
водорослей.
С
продвижением в высокие широты эти пики
все более сближаются
друг
с другом во времени. В высокоарктических
водах из-за крат-
кости
вегетационного периода наблюдается
только один пик чис-
ленности
водорослей.
В
морях четко прослеживается зависимость
численности водо-
рослей
от вертикальной устойчивости водных
масс. С одной сторо-
ны,
перемешивание водных масс в вертикальном
направлении бла-
гоприятно
для развития водорослей, так как
обеспечивает попол-
нение
поверхностных слоев биогенами за счет
их выноса из глу-
бин.
С другой стороны, вертикальные токи
воды могут заносить
водоросли
на такие глубины, где света для них
недостаточно. Чем
толще
слой воды, в котором рассредоточены
водоросли, тем меньше
света
они могут утилизировать, поскольку
часть его, увеличиваю-
щаяся
с продвижением вглубь, поглощается
водой. Помимо этого,
жизнедеятельность
вертикально перемещаемых водорослей
угне-
тается
и в другом отношении: они не могут
приобрести температур-
ных
и световых адаптаций, повышающих
эффективность фотосин-
теза.
Такие свойства вырабатываются только
в условиях четкой
вертикальной
стратификации водной массы, только в
присутствии
слоя
плотностного скачка. Например, из-за
высокой перемешивае-
мости
вод в зимнее время у берегов Англии
фитопланктон беден,
хотя
света вполне достаточно для развития
водорослей. По сход-
ным
причинам в прибрежных водах цветение
фитопланктона начи-
нается
раньше, поскольку здесь быстрее
устанавливается плотност-
ная
стратификация. Прибрежные воды,
опресняемые стоком с су-
ши
и благодаря этому становящиеся менее
плотными, по мере
таяния
снегов тонким слоем разливаются по
поверхности более со-
леных
вод. В результате образуется плотностная
стратификация,
область
распространения которой расширяется
в направлении от
прибрежья
в открытое море, соответственно чему
в цветение после-
довательно
включаются все новые, более удаленные
от берега
участки
моря.
279
Растеканием
талой воды по поверхности соленой и
образова-
нием
плотностной стратификации объясняется
интенсивное разви-
тие
водорослей у кромки тающего льда,
которая является как бы
«ложным
берегом». Нарушением плотностной
стратификации воды
можно
объяснить и угнетающее действие штормов
на развитие фи-
топланктона.
Сезонная
динамика зоопланктона.
Сезонные коле-
бания
биомассы и численности популяций
планктонных животных
связаны
главным образом с периодичностью
массового появления
в
водоемах водорослей и других растений,
прямо или косвенно слу-
жащих
пищей зоопланктону. Колебания термики
представляют со-
бой
второй важный фактор, определяющий
сезонность изменения
численности
планктонных животных. Третий важнейший
фактор
динамики
зоопланктона — сезонность его выедания
рыбами и дру-
гими
животными. Так как основная часть
планктонных животных
растительноядна,
зоопланктон обогащается количественно
только
после
массового появления водорослей, причем
с некоторым опоз-
данием.
В самом зоопланктоне растительноядные
формы появляют-
ся
в массовом количестве раньше
зоопланктофагов, развитие кото-
рых
обусловливает повышение численности
хищников, живущих за
счет
плотоядных планктонных животных.
Так
как с продвижением в высокие широты
продолжительность
вегетации
водорослей снижается, то соответственно
сокращаются
и
периоды высокой численности зоопланктона.
Параллельно этому
возрастают
сезонные колебания количества
зоопланктона, которые
сравнительно
невелики в низких широтах и более резки
в высоких.
В
некоторых случаях массовое появление
водорослей сопровож-
дается
угнетением зоопланктона. Например, во
время цветения
воды
в Байкале снижается численность рачка
Ер1$сНига
Ьшса1еп-
818,
наибольшее богатство зоопланктона
здесь наблюдается после
некоторого
уменьшения количества водорослей. В
прудах неодно-
кратно
наблюдалось снижение численности
веслоногих рачков и
коловраток
в периоды массового появления синезеленой
АрНатго-
тепоп.
В условиях эксперимента 'Прослежено
отрицательное влия-
ние
СМогеИа
ругепоШоза
на рачка ОарНша
тацпа\
гибель послед-
него
наблюдалась в культуре синезеленой
АпаЬаепа
Vа^^аЫ^^8.
По-
видимому,
жизнедеятельность многих представителей
зоопланкто-
на
угнетают высокие концентрации некоторых
метаболитов, созда-
ющиеся
во время массового появления водорослей.
Богатство
зоопланктона резко возрастает в периоды
размноже-
ния
донных животных за счет появления их
пелагических личинок
через
некоторое время после весенней вспышки
фитопланктона.
Личинки
живут в толще воды от нескольких недель
в водоемах
высоких
широт до многих месяцев в тропических
водах. После
достижения
максимума численность зоопланктонов
начинает сни-
жаться
в результате их выедания и отмирания.
Сезонная
динамика зообентоса.
Сезонные колеба-
ния
численности и биомассы донных животных
в первую очередь
зависят
от особенностей их размножения, роста
и выедания, а так-
280
же
от ряда абиотических факторов, в
частности термики водоемов.
Последняя
определяет время размножения донных
животных как
непосредственно,
так и косвенно через появление
водорослей, кото-
рыми
питаются пелагические личинки бентосных
организмов.
В
континентальных водоемах резкие
колебания численности и
биомассы
популяций донных животных могут
обусловливаться мас-
совым
вылетом насекомых. В прудах и озерах,
где на долю личи-
нок
насекомых нередко приходится более
половины всей биомассы
бентоса,
его обеднение во время вылета имаго
часто носит опусто-
шительный
характер. В зависимости от количества
генераций у
массовых
форм насекомых резкие снижения биомассы
бентоса в
континентальных
водоемах происходят от одного до 3—4
раз в год.
Например,
у большинства личинок хирономид в
Волге у Саратова
наблюдаются
три минимума численности в бентосе,
что соответст-
вует
такому же числу массовых вылетов
комаров. Вылет хироно-
мид
иногда столь дружен, что за несколько
недель их личинки поч-
ти
совершенно исчезают из бентоса.
Годовая
динамика. Нередко в водоемах или их
отдельных уча-
стках
на протяжении нескольких лет происходят
циклические из-
менения
численности и биомассы популяций.
Обычно они связаны
с
периодическими колебаниями интенсивности
солнечной радиации,
многолетними
колебаниями уровня водоемов, с изменением
их со-
лености,
кислородного режима и некоторых других
свойств. Как
правило,
периодичность годовых колебаний
численности в связи с
изменениями
солнечной активности — 9—12 лет.
Например, раз в
12
лет теплое течение Эль-Ниньо достигает
берегов Перу, от-
тесняя
холодное Перуанское течение, и в это
время из-за недостат-
ка
кислорода и заметного повышения
температуры воды катастро-
фически
гибнут планктонные животные и рыбы.
В годы повышен-
ной
инсоляции в Северном Каспии и Азовском
море в связи с со-
кращением
стока равнинных рек и уменьшением
количества при-
носимых
ими питательных солей условия
воспроизводства популя-
ций
гидробионтов ухудшаются. Наоборот, в
Южном Каспии и
Аральском
море, куда впадают реки с ледниковым
питанием, по-
ступление
биогенов в годы с повышенной инсоляцией
увеличивает-
ся
и условия существования популяций
многих гидробионтов улуч-
шаются.
Нередко
годовые колебания численности
гидробионтов не свя-
заны
с геофизическими циклами. К ним, в
частности, относятся те
периодические
изменения, которые возникают в результате
взаимо-
действия
популяций жертвы и хищника. Когда
численность жертвы
достигает
высоких величин, условия питания
хищников улучшают-
ся
и их в водоеме становится больше. Это
увеличение ведет к уси-
лению
выедания жертвы, ее популяции становятся
малочисленнее
и
соответственно ухудшаются условия
воспроизводства хищника,
его
численность снова снижается. Подобная
динамика численности
наблюдается
и в популяциях промысловых объектов
как следствие
вылова.
Когда их концентрация велика, добыча
делается особенно
выгодной
и объем вылова возрастает, обусловливая
резкое сни-
281
Годы Годы
Рис.
48. Годовые колебания численности
некоторых экологически близких
бен-
тосных
форм в датских водах (по Мооге, 1958).
Пунктир — формы с длительно-
живущей
пелагической личинкой, сплошная линия
— пелагических личинок нет или
они
существуют очень короткое время; Л —
СиНеИиз
реИисШия
(1)
и Ыиси1а
пШго
(2);
Б
— А1Ьа
а1Ьа
(3) и Масота
са1сагеа (4);
В
— СогЬи1а
ц1ЬЬа (5)
и А;иси1а
пШАа
(6)
жение
численности промыслового стада
животных. В результате
промысел
прекращается и начинается восстановление
стада.
Годовые
колебания численности резче выражены
у форм с ко-
ротким
жизненным циклом. У видов с длительно
существующими
пелагическими
личинками численность популяций менее
устойчива,
чем
у организмов с прямым развитием или с
краткой пелагической
стадией
развития (рис. 48). Первые формы,
характеризующиеся
высокой
плодовитостью, но вместе с тем и малой
жизнестойкостью
зародышей,
дают высокий .подъем численности в
годы с благо-
приятными
условиями и резкое снижение в годы,
неблагоприятные
для
размножения.
Непериодические
изменения. В ряде случаев численность
и био-
масса
популяций испытывают существенные
изменения, не сопро-
вождающиеся
возвратом к исходному состоянию. Такие
неперио-
дические
сдвиги прежде всего обусловливаются
ациклическими из-
менениями
режима водоемов, в частности, вызываемые
деятель-
ностью
человека, — гидростроительством,
водопотреблением и во-
допользованием,
сбросом сточных вод и др. В результате
зарегули-
рования
стока рек и соответствующего изменения
их гидрологиче-
ского
режима численность популяции одних
гидробионтов возрас-
тает,
другие становятся малочисленнее или
исчезают вовсе.
282