

2.2Окисление жира
Образовавшиеся в результате гидролиза триацилглицерола (см. рис. 7) глицерин и жирные кислоты вовлекаются в дальнейший путь распада – окисление.
2.2.1Окисление глицерина
Глицерин вовлекается в цитоплазме клетке в процесс гликолиза.
Сначала глицерин при участии фермента глицерофосфаткиназы (трансфераза) превращается в -глицеролфосфат. Последний под действием НАД+-зависимой -глицеролфосфатдегидрогеназы (оксидоредуктаза) превращается в фосфодиоксиацетон:
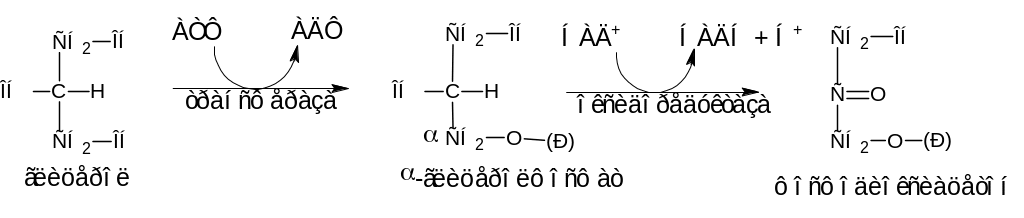
Фосфодиоксиацетон, являясь обычным метаболитом гликолиза, изомеризуется в 3-фосфоглицериновый альдегид и включается в метаболический путь (гликолиз) до ПВК. Пировиноградная кислота, превращаясь в ацетил-КоА, полностью окисляется в цикле Кребса до СО2, Н2О и выделением АТФ.
2.2.2Окисление жирных кислот
Ф.Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях живых организмов происходит в -положении. В результате от молекулы жирной кислоты отщепляются двууглеродные фрагменты со стороны карбоксильной группы. Процесс -окисления жирных кислот складывается из следующих этапов:
Активация жирных кислот. Подобно первой стадии гликолиза сахаров перед -окислением жирные кислоты подвергаются активации. Эта реакция протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима А (НS-КоА) и ионов Mg2+. Реакция катализируется ацил-КоА-синтетазой. В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты, АМФ и РРi (Н4Р2О7).

Транспорт жирных кислот внутрь митохондрий. Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и протекает их окисление, переносчиком активированных жирных кислот через внутреннюю митохондриальную мембрану служит карнитин (-триметиламино--оксибути-рат):

После прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепление ацилкарнитина при участии НS-КоА и митохондриальной карнитин-ацилтрансферазы:
Далее ацил-КоА в митохондрии подвергается -окислению.
В результате этого процесса происходит последовательное отщепление от карбоксильного конца углеродной цепи жирной кислоты двууглеродных фрагментов в форме ацетила-КоА и соответствующее укорачивание цепи жирной кислоты:

Жирная кислота с четным числом атомов углерода
В матриксе митохондрии -окисление ацил-КоА осуществляется в результате повторяющейся последовательности четырех реакций (рис.8).
1) окисления с участием ацил-КоА-дегидрогеназы (ФАД-зависимой дегидрогеназы);
2) гидратации, катализируемой еноил-КоА-гидратазой;
3) второго окисления под действием 3-гидроксиацил-КоА-дегидрогеназы (НАД-зависимой дегидрогеназы);
4) тиолазной с участием ацетил-КоА-ацилтрансферазы.
Совокупность этих четырех последовательностей реакций составляет один цикл -окисления жирной кислоты (см. рис. 8).
Образовавшийся ацетил-КоА подвергается окислению в цикле Кребса, а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь -окисления вплоть до образования бутирил-КоА (4-углеродное соединение), который на последнем этапе -окисления распадается на две молекулы ацетил-КоА.
При окислении жирной кислоты, содержащей n углеродных атомов, происходит n2-1 циклов -окисления (т.е. на один цикл меньше, чем n2, так как при окислении бутирил-КоА сразу происходит образование двух молекул ацетил-КоА) и всего получится n2 молекул ацетил-КоА.
Например, при окислении пальмитиновой кислоты (С16) повторяется 162-1=7 циклов -окисления и образуется 162=8 молекул ацетил-КоА.
Баланс энергии. При каждом цикле -окисления образуется одна молекула ФАДН2 (см. рис. 8; реакция 1) и одна молекула НАДН+Н+(реакция 3). Последние в процессе окисления в дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 – 2 молекулы АТФ и НАДН+Н+ – 3 молекулы АТФ, т.е. в сумме за один цикл образуется 5 молекул АТФ. При окислении пальмитиновой кислоты образуется 57=35 молекул АТФ. В процессе -окисления пальмитиновой кислоты образуется 8 молекул ацетил-КоА, каждая из которых, «сгорая» в цикле Кребса, дает 12 молекул АТФ, а 8 молекул дадут 128=96 молекул АТФ.
Таким образом, всего при полном окислении пальмитиновой кислоты образуется 35+96=131 молекула АТФ. С учетом одной молекулы АТФ, потраченной в самом начале на активацию жирной кислоты, общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты составит 131–1=130 молекул АТФ.
Однако образовавшийся в результате -окисления жирных кислот ацетил-КоА может не только окисляться до СО2, Н2О, АТФ, вступая в цикл Кребса, но использоваться на синтез холестерина , а также углеводов в глиоксилатном цикле.
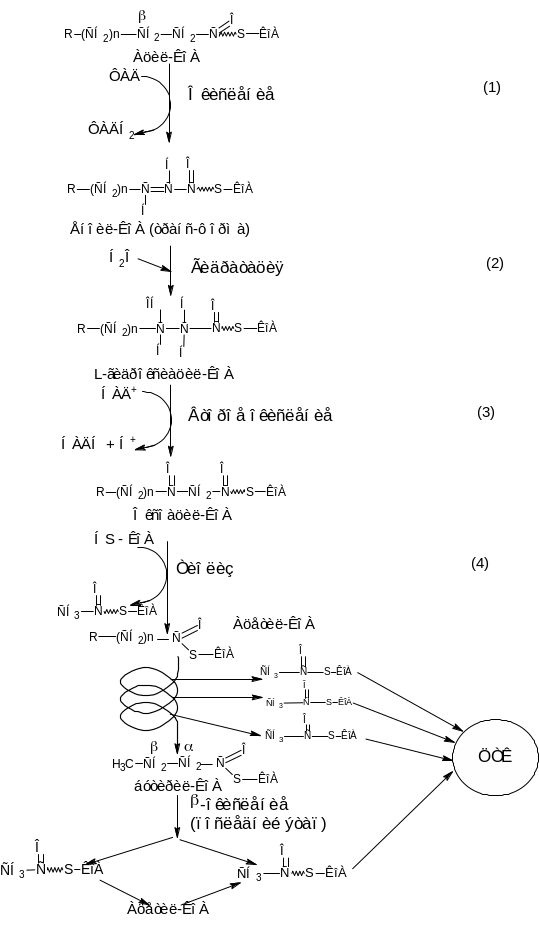
Рисунок 8 – Схема -окисления жирной кислоты
Глиоксилатный цикл специфичен только для растений и бактерий, у животных организмов он отсутствует. Данный процесс синтеза углеводов из жиров подробно описан в методическом указании /8/.
Окисление ненасыщенных жирных кислот
Олеиновая, линолевая и линоленовая кислоты, являющиеся важнейшими компонентами ацилглицеролов, также подвергаются -окислению. Окисление этих ненасыщенных жирных кислот, в принципе, происходит так же, как и окисление насыщенных жирных кислот, но с некоторыми особенностями.
При окислении олеиновой кислоты (содержащей одну двойную связь) в результате трех циклов -окисления (рис. 8) образуется 3 молекулы ацетил-КоА и 12-углеродная ненасыщенная жирная кислота с цис-двойной связью между 3-им и 4-ым атомами углерода.
Дело в том, что образовавшаяся 12-углеродная ненасыщенная жирная кислота должна снова включиться в путь -окисления, но двойные связи природных ненасыщенных жирных кислот имеют цис-конфигурацию, а при -окислении насыщенных жирных кислот двойные связи имеют транс-конфигурацию. Кроме того, в результате последовательного удаления трех двууглеродных фрагментов (ацетил-КоА) до первой двойной связи дает 3,4– ацил-КоА (расположение двойной связи между 3 и 4 атомами углерода), а не 2,3– ацил-КоА, который является промежуточным продуктом при -окислении насыщенных жирных кислот. Поэтому в тканях существует фермент, который осуществляет перемещение двойной связи из положения 3-4 в положение 2-3, а также изменяет конфигурацию двойной связи из цис- в транс-положение.
Этот
фермент получил название: 3,4-цис
→ 2,3
-транс-еноил-КоА-изомеразы
3,4-цис
→ 2,3
-транс-еноил-КоА-изомеразы
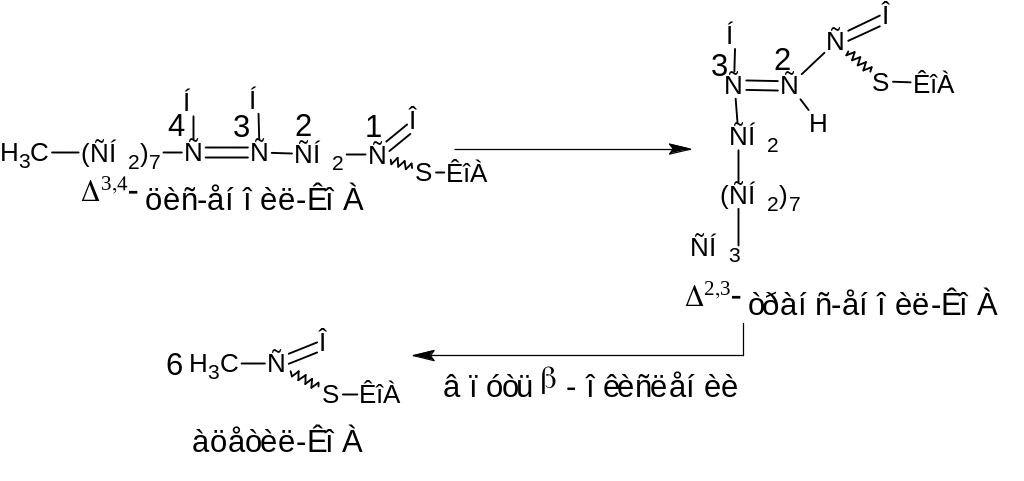
Образовавшийся 2,3-транс-еноил-КоА включается в путь -окисления (см рис.8) в стадию гидратации (2).
В результате полного окисления олеиновой кислоты образуется 9 (6+3) молекул ацетил-КоА, которые включаются в цикл Кребса и «сгорают» до СО2, Н2О с образованием энергии.
Окисление жирных кислот с нечетным числом углеродных атомов
Основная масса природных липидов содержит жирные кислоты с четным числом углеродных атомов, однако в липидах многих растений и некоторых морских организмов присутствуют жирные кислоты с нечетным числом атомов углерода.
Установлено, что жирные кислоты с нечетным числом углеродных атомов окисляются таким же образом, как и жирные кислоты с четным числом углеродных атомов, с той лишь разницей, что на последнем этапе расщепления (-окисления) образуется одна молекула пропионил-КоА и одна молекула ацетил-КоА, а не 2 молекулы ацетил-КоА (рис. 8)? как в случае -окисления жирных кислот с четным числом углеродных атомов:
Рисунок 9 – Схема -окисления жирных кислот с нечетным числом
углеродных атомов
Окисление пропионил-КоА
Пропионил-КоА, являющийся одним из конечных продуктов -окисления жирных кислот с нечетным числом углеродных атомов, превращается в сукцинил-КоА путем двух последовательных реакций.
Реакция карбоксилирования пропионила:

2. Реакция внутримолекулярой перегруппировки:

Сукцинил-КоА далее утилизируется, поступая в цикл Кребса:

Но при прохождении всех этих реакций сукцинил-КоА до ЩУК по циклу Кребса не происходит полного окисления до СО2 и Н2О. Для этого образовавшийся оксалоацетат через ряд последовательных реакций превращается в ацетил-КоА, который, поступая в цикл Кребса, полностью «сгорает» до СО2 , Н2О и выделением энергии.
Превращение оксалоацетата в ацетил-КоА
1) Образовавшийся оксалоацетат локализован в митохондрии. Мембрана митохондрий непроницаема для образовавшегося оксалоацетата. Последний здесь же в митохондрии восстанавливается в яблочную кислоту:

Реакция протекает при участии митохондриальной НАД-зависимой малатдегидрогеназы. Образовавшаяся яблочная кислота легко выходит из митохондрии в цитозоль клетки и вновь окисляется в щавелевоуксусную кислоту при участии цитоплазматической НАД-зависимой малатдегидрогеназы:

2) Превращение оксалоацетата в фосфоенолпируват происходит в цитоплазме клетки:

3) Образовавшийся фосфоенолпируват (ФЕП) по пути гликолиза превращается в ПВК, а пируват в результате окислительного декарбоксилирования в ацетил-КоА, который поступая в цикл Кребса, полностью окисляется до СО2 и Н2О, с выделением энергии.
С учетом вышесказанного видно, что образовавшийся пропионил, в конечном счете, должен превратиться в ацетил-КоА и окислиться в цикле Кребса. Таким образом, все жирные кислоты в результате -окисления превращаются в ацетил-КоА, который «сгорает» в цикле Кребса.