

Превращение энергии в процессе фотосинтеза. Характеристика 4 стадий фотосинтеза: суть энергетических превращений, скорость реакций, химизм, локализация внутри хлоропластов.
В настоящее время установлено, что фотосинтез протекает в две стадии: световую и темновую. Световая стадия – это процесс использования света для расщепления воды; при этом выделяется кислород и образуются богатые энергией соединения.
Темновая стадия включает группу реакций, в которых используются высокоэнергетические продукты световой стадии для восстановления СО2 до простого сахара, т.е. для ассимиляции углерода. Поэтому темновую стадию называют также стадией синтеза. Термин «темновая стадия» означает лишь то, что свет в ней непосредственно не участвует.
У растений реакции фотосинтеза протекают в хлоропластах - специальных энергопреобразующих органеллах, которые за счет энергии солнечного света обеспечивают синтез углеводов из углекислого газа и воды. В хлоропластах источником энергии для синтеза углеводов являются молекулы аденозинтрифосфорной кислоты (АТР) и восстановленного никотинамидадениндинуклеотидфосфата (NADPH).
Световая энергия поглощается пигментами. У всех растений, осуществляющих фотосинтез, имеются различные формы хлорофилла, и, вероятно, во всех содержатся каротиноиды, окрашенные обычно в желтые тона. В высших растениях содержатся хлорофилл а (С55Н72О5N4Mg) и хлорофилл b (C55H70O6N4Mg), а также четыре основных каротиноида: b-каротин (С40Н56), лютеин (С40Н55О2), виолаксантин и неоксантин. Такое разнообразие пигментов обеспечивает широкий спектр поглощения видимого света, поскольку каждый из них «настроен» на свою область спектра.
Фотосистемы. Хлорофилл и каротиноиды, погруженные в тилакоидные мембраны хлоропластов, собраны в функциональные единицы – фотосистемы, каждая из которых содержит примерно 250 молекул пигментов. Устройство фотосистемы таково, что из всех этих молекул, способных поглощать свет, только одна особым образом расположенная молекула хлорофилла а может использовать его энергию в фотохимических реакциях – она является реакционным центром фотосистемы. Остальные молекулы пигментов, поглощая свет, передают его энергию на реакционный центр; эти светособирающие молекулы называют антенными. Существует два типа фотосистем. В фотосистеме I специфическая молекула хлорофилла а, составляющая реакционный центр, имеет оптимум поглощения при длине световой волны 700 нм (обозначается P700; P – пигмент), а в фотосистеме II – при 680 нм (P680). Обычно обе фотосистемы работают синхронно и (на свету) непрерывно, хотя фотосистема I может работать и отдельно.
Световая фаза
Эта фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это приводит к распаду или фотолизу воды:
Н2О + Qсвета → Н+ + ОН—.
Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы •ОН:
ОН— → •ОН + е—.
Радикалы •ОН объединяются, образуя воду и свободный кислород:
4НО• → 2Н2О + О2.
Кислород при этом удаляется во внешнюю среду, а протоны накапливаются внутри тилакоида в «протонном резервуаре». В результате мембрана тилакоида с одной стороны за счет Н+ заряжается положительно, с другой за счет электронов — отрицательно. Когда разность потенциалов между наружной и внутренней сторонами мембраны тилакоида достигает 200 мВ, протоны проталкиваются через каналы АТФ-синтетазы и происходит фосфорилирование АДФ до АТФ; атомарный водород идет на восстановление специфического переносчика НАДФ+ (никотинамидадениндинуклеотидфосфат) до НАДФ·Н2:
2Н+ + 2е— + НАДФ → НАДФ·Н2.
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.
Темновая фаза
Эта фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
Первая реакция в этой цепочке — фиксация углекислого газа; акцептором углекислого газа является пятиуглеродный сахаррибулозобифосфат (РиБФ); катализирует реакцию фермент рибулозобифосфат-карбоксилаза (РиБФ-карбоксилаза). В результате карбоксилирования рибулозобисфосфата образуется неустойчивое шестиуглеродное соединение, которое сразу же распадается на две молекулы фосфоглицериновой кислоты (ФГК). Затем происходит цикл реакций, в которых через ряд промежуточных продуктов фосфоглицериновая кислота преобразуется в глюкозу. В этих реакциях используются энергии АТФ и НАДФ·Н2, образованных в световую фазу; цикл этих реакций получил название «цикл Кальвина»:
6СО2 + 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды. В настоящее время различают два типа фотосинтеза: С3- и С4-фотосинтез.
С3-фотосинтез
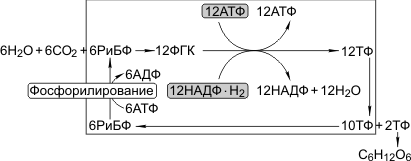
Это тип фотосинтеза, при котором первым продуктом являются трехуглеродные (С3) соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза (М. Кальвин). Именно С3-фотосинтез описан как «Темновая фаза». Характерные особенности С3-фотосинтеза: 1) акцептором углекислого газа является РиБФ, 2) реакцию карбоксилирования РиБФ катализирует РиБФ-карбоксилаза, 3) в результате карбоксилирования РиБФ образуется шестиуглеродное соединение, которое распадается на две ФГК. ФГК восстанавливается до триозофосфатов (ТФ). Часть ТФ идет на регенерацию РиБФ, часть превращается в глюкозу.
С4-фотосинтез
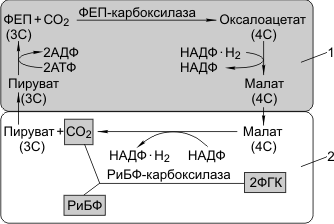
С4-фотосинтез — фотосинтез, при котором первым продуктом являются четырехуглеродные (С4) соединения. В 1965 году было установлено, что у некоторых растений (сахарный тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются четырехуглеродные кислоты. Такие растения назвали С4-растениями. В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый газ. Путь превращений углерода в С4-растениях стали называть путем Хэтча-Слэка.
Для С4-растений характерно особое анатомическое строение листа. Все проводящие пучки окружены двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки. Углекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват (ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С). Процесс катализируется ФЕП-карбоксилазой. В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2 и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла много гран, где активно идут реакции световой фазы. В хлоропластах клеток обкладки идут реакции темновой фазы.
Оксалоацетат (4С) превращается в малат, который через плазмодесмы транспортируется в клетки обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата, СО2 и НАДФ·Н2.
Пируват возвращается в клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
|
|
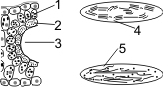
Строение С4-растений: 1 — наружный слой — клетки мезофилла; 2 — внутренний слой — клетки обкладки; 3 — «Кранц-анатомия»; 4, 5 — хлоропласты; 4 — многочисленные граны, крахмала мало; 5 — немногочисленные граны, крахмала много. |
С4-фотосинтез: 1 — клетка мезофилла; 2 — клетка обкладки проводящего пучка. |
|
|
Строение фотосинтезирующих мембран хлоропластов, методы их исследования: метод реплик с замороженных сколов, дифференциального центрифугирования. Фотосистема 1 и фотосистема 2 как компоненты ЭТЦ фотосинтеза.
Структурным компонентом внутренних мембран хлоропластов является
тилакоидная мембранная система, окружённая внутренней гомогенной средой,
стромой. Тилакоидная мембрана представляет собой липидный бислой (Staehelin et al., 1996) и содержит мульти-ферментные комплексы, осуществляющие разделение зарядов и перенос электрона через мембрану – комплексы фотосистемы I, фотосистемы II, цитохромный b6f комплекс, АТФ-синтазу. В мембранах тилакоидов локализованы пигменты и другие компоненты,
связанные с поглощением и использованием энергии света, биохимические
системы синтеза и превращения углеводов функционируют в строме хлоропластов. Тилакоидная мембрана имеет сложную пространственную организацию. Часть мембран формируют стопки (граны), соединенные между собой перемычками (стромальными ламеллами). Трансмембранные белковые комплексы распределены в мембране тилакоида неоднородно. В гранальных участках тилакоидной мембраны находятся комплексы ФС2. Комплексы ФС1 и ATФ-синтазы в основном встречаются в стромальных и краевых, выдающихся в строму участках тилакоидной мембраны. Цитохромный b6f комплекс распределен в мембране однородно.
Начальной стадией фотосинтеза является поглощение света молекулами
пигментов, входящих в состав светособирающих или антенных комплексов, и
передача энергии возбуждённого состояния на реакционные центры. Ассоциированные в липидной мембране светособирающие комплексы состоят из
белков, хлорофиллов и каротеноидов, и характеризуются высокой степенью
организации. Важнейшая роль светособирающих белков состоит в регуляции
фотосинтеза и в обеспечении фотозащитной функции.
Характерной особенностью тилакоидных мембран является их отрицательный поверхностный заряд. На наружной поверхности мембран плотность зарядов выше. Поверхностный заряд играет важную роль в процессе формирования гран. Катионы, экранируя отрицательные заряды на поверхности мембран, снижают силы их отталкивания, что ведет к слипанию мембран и образованию стопки тилакоидов – граны (процесс стэкинга).
метод электронной микроскопии «замораживание-скалывание» - дает возможность изучать внутреннее строение клеточных мембран. Клетки замораживают при температуре жидкого азота (-196°С) в присутствии криопротектора (антифриза) во избежание искажений за счет образования кристаллов льда. Замороженный блок затем раскалывают лезвием ножа. когда плоскость среза проходит через мембрану, последняя раскалывается преимущественно по своей срединной области и расщепляется на две половинки. В результате на образовавшихся плоскостях скола обнажается внутренняя область мембраны.Образующуюся поверхность скола оттеняют платиной, органический материал удаляют и изучают полученные реплики в электронном микроскопе. Такие реплики усеяны небольшими выпячиваниями - внутримембранными частицами, которые представляют собой крупные мембранные белки.
Для того чтобы изучить состав и функции тех или иных клеток, применяют метод дифференциального центрифугирования. Он основан на том, что различные клеточные органеллы и включения имеют различную плотность. При очень быстром вращении в специальном приборе - ультрацентрифуге - органеллы тонко измельченных клеток выпадают в осадок из раствора, располагаясь слоями в соответствии со своей плотностью: более плотные компоненты осаждаются при более низких скоростях центрифугирования, а менее плотные - при более высоких скоростях. Эти слои разделяют и изучают отдельно.
Фотохимический этап фотосинтеза включает ряд последовательно протекающих процессов, локализованных в тилакоидных мембранах. Пигменты, специфически связанные с белками фотосинтетических мембран, и другие компоненты, необходимые для протекания реакций поглощения света и электронного транспорта, образуют надмолекулярные комплексы — фотосистему I (ФС I) и фотосистему II (ФС II). В составе каждой фотосистемы различают: реакционный центр, в котором происходят очень быстрые реакции первичного разделения зарядов; комплекс компонентов, по которым передается электрон от реакционного центра, и последний окисляется (электронтранспортная цепь); комплекс компонентов, за счет работы которых происходит фотоокисление воды и восстановление реакционного центра.
Существует два типа фотосистем. В фотосистеме I специфическая молекула хлорофилла а, составляющая реакционный центр, имеет оптимум поглощения при длине световой волны 700 нм (обозначается P700; P – пигмент), а в фотосистеме II – при 680 нм (P680). Обычно обе фотосистемы работают синхронно и (на свету) непрерывно, хотя фотосистема I может работать и отдельно.
Первый этап сложного преобразования электромагнитного излучения (света) в свободную энергию химических связей включает поглощение фотонов светособирающими комплексами (антеннами), связанными с ФС I и ФС II (ССКI и ССКII, соответственно). Затем энергия возбуждения мигрирует по пигментам антенны (от более коротковолновых форм хлорофилла к более длинноволновым) и захватывается ловушкой — специализированным реакционным центром, который расположен в центре комплекса. Реакционные центры образованы самыми длинноволновыми формами хлорофилла а [с максимумом поглощения 700 нм (Р700) в ФС I и 680 нм (Р680) в ФС II]. Возбужденные Р700* и Р680* — очень сильные восстановители и быстро передают электрон на близко расположенную молекулу акцептора, а сами при этом окисляются. Эти реакции первичного разделения зарядов, происходящие в реакционных центрах ФС I и ФС II, являются единственными, в которых действительно происходит превращение энергии кванта света в химическую энергию. Дальнейший транспорт электронов, препятствующий рекомбинации зарядов, осуществляется по градиенту электрохимического потенциала компонентов электрон-транспортной цепи фотосинтеза.