

Причины высокой каталитической активности.
Фермент поддерживает микроокружение субстрата в активном центре в состоянии отличном от его состояния в водной среде. Располагает реагирующие атомы в правильной ориентации и на необходимом расстоянии друг от друга, чтобы обеспечить оптимальное протекание реакции. За счет кооперативного взаимодействия субстрата и нескольких остатков аминокислот в активном центре фермент снижает энергию активации данной реакции.
Субстратная специфичность
Структура активного центра фермента комплементарна структуре субстрата, т.е. соответствует ему по 1) форме, 2) размерам и 3) способности взаимодействовать. Это является причинами высокой специфичности ферментов. Первоначально модель активного центра, предложенная Э.Фишером, трактовала взаимодействие субстрата и фермента по аналогии с системой «ключ-замок» – модель «жесткой матрицы». Однако эта модель объясняла лишь абсолютную субстратную специфичность.
Кошланд предложил модель индуцированного соответствия. Главная черта этой модели – гибкость каталитического центра. В модели Фишера каталитический центр считается заранее подготовленным под форму молекулы-субстрата. В модели Кошланда субстрат индуцирует конформационные изменения фермента, и лишь в результате этих аминокислотные остатки и другие группы фермента принимают пространственную ориентацию, необходимую для связи с субстратом и катализа. Эта модель позволяет объяснить относительную специфичность фермента.
Каждый фермент катализирует не любые из всех возможных путей превращения субстрата, а какое-либо одно. Это свойство называется специфичностью пути превращения.
Кинетика ферментативных реакций
Зависимость скорости ферментативной реакции от концентрации субстрата. Уравнение Михаэлиса-Ментен.Любую ферментативную реакцию схематично можно описать следующим образом:
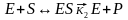
Поведение многих ферментов при изменении концентрации субстрата описывает уравнение Михаэлиса-Ментен:
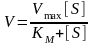 ,
где
,
где
 -
скорость ферментативной реакции,
-
скорость ферментативной реакции,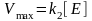 -
максимальная скорость ферментативной
реакции при полном насыщении фермента
субстратом,
-
максимальная скорость ферментативной
реакции при полном насыщении фермента
субстратом,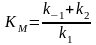 – константа Михаэлиса,
– константа Михаэлиса,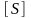 –
концентрация субстрата.
–
концентрация субстрата.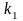 -
константа образования фермент-субстратного
комплекса ES,
-
константа образования фермент-субстратного
комплекса ES, -
константа диссоциации фермент-субстратного
комплекса ES.
-
константа диссоциации фермент-субстратного
комплекса ES.
В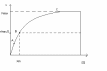 лияние
концентрации субстрата на скорость
ферментативной реакции, описываемое
этим уравнением, можно изобразить
графически:
лияние
концентрации субстрата на скорость
ферментативной реакции, описываемое
этим уравнением, можно изобразить
графически:
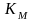 равна
концентрации субстрата при скорости
ферментативной реакции, равной половине
максимальной скорости и характеризует
сродство данного фермента к тому или
иному субстрату. Некоторые ферменты
требуют высокой концентрации субстрата
для достижения скорости, равной
максимальной, другие (например,
гексокиназа) достигают
равна
концентрации субстрата при скорости
ферментативной реакции, равной половине
максимальной скорости и характеризует
сродство данного фермента к тому или
иному субстрату. Некоторые ферменты
требуют высокой концентрации субстрата
для достижения скорости, равной
максимальной, другие (например,
гексокиназа) достигают
 при очень низкой концентрации субстрата.
при очень низкой концентрации субстрата.
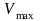 - скорость реакции при 250С,
оптимальном значении рН, полном насыщении
фермента субстратом.
- скорость реакции при 250С,
оптимальном значении рН, полном насыщении
фермента субстратом. -
число оборотов
фермента,
т.е. число молекул субстрата, которое
превращает одна молекула фермента за
1 минуту.
-
число оборотов
фермента,
т.е. число молекул субстрата, которое
превращает одна молекула фермента за
1 минуту.
Например:
карбоксиангидраза 36000000
-амилаза 1100000
фосфоглюкомутаза 1240