

Принцип корреляции (ж. Кювье)
По Кювье, в организме как целом, части которого связаны функционально, изменение любого отдельного органа затрагивает другие органы. По И. И. Шмальгаузену (1938) понятие корреляции, как его понимал Ж. Кювье, теряет определенность и становится не нужным. Следует говорить о корреляциях при наличии ясной связи между чем-либо. Выделяют генетические корреляции (как результат плейотропного влияния гена), морфогенетические (как результат индукций), функциональные (как результат функционального взаимодействия структур).
Принцип независимой эволюции отдельных морфологических признаков (г. Осборн, 1910)
Согласно этому принципу, морфологические структуры организма, функционально не связанные друг с другом, могут в процессе эволюции изменяться относительно независимо. Принцип Осборна объясняет мозаичность переходных форм и является очень важным для сравнительной анатомии в
методическом плане, т.к. дает основание для изучения
преобразований в эволюции отдельных органов.
Принцип дифференциации и интеграции
По И.И. Шмальгаузену (1938), это основной принцип развития, связанный с преобразованием органов. В основе дифференциации лежит морфологическое обособление частей, при этом выделенная из целого часть выполняет одну определенную функцию (чаще несколько функций из их ряда). Последнее объясняет, почему дифференциация обязательно сопровождается интеграцией – соподчинением частей целому, Но уже на другом уровне.
Дифференциация почти никогда не приводит к тому, что часть выполняет лишь одну функцию. Органы высших животных мультифункциональны. У каждого из них есть характерная функция – главная и второстепенные – побочные функции.
Принцип взаимосвязи форм и функций
В процессах филогенетического преобразования органов форма и функция неразрывно связаны, поэтому любое подобное преобразование есть морфофункциональное (А. Н. Северцов).
Способы филогенетических преобразований органов
Смена функций. При изменении условий главная функция теряет свое значение, а одна из второстепенных
функций становится главной.
Расширение функций. Приобретение структурой новых функций может сопровождаться дифференциацией органа.
Сокращение функций. Уменьшение числа функций часто наблюдается при специализации, когда прогрессирует лишь одна (или несколько) функция из их ряда.
Разделение функций. Наблюдается при дифференциации целого.
Субституция органов. В процессе прогрессивного развития часто наблюдается замена одного органа на аналогичный, выполняющий ту же функцию.
Интенсификация функций. Усовершенствованное выполнение какой-либо функции органом в результате его морфологических изменений.
СРАВНИТЕЛЬНАЯ АНАТОМИЯ ПОКРОВОВ Функциональное значение покровов
Покровы хордовых выполняют многообразные функции, связанные с обеспечением разнообразных форм взаимосвязи организма с окружающей средой:
Отграничительная функция – покровы отграничивают внутреннюю среду организма от внешней среды.
Защитная функция – покровы обеспечивают защиту организма от механических и химических воздействий, проникновения болезнетворных организмов.
Обменная функция – через покровы осуществляется обмен различными веществами между организмом и внешней средой, в частности водно-солевой обмен, газообмен.
Рецепторная функция – специализированные клетки покровов способны к специфическому восприятию различных раздражений со стороны внешней среды.
5. Функция терморегуляции – через покровы осуществляется теплообмен между организмом и внешней средой.
Наряду с этими функциями покровы различных хордовых животных могут выполнять некоторые специальные функции, например, образование пахучих и ядовитых веществ, запасание питательных веществ, участие в формировании скелетных структур и др.
Общая характеристика покровов хордовых
В строении покровов хордовых повторяется общая схема, характерная для покровов вторичноротых животных. Покровы всегда двуслойны: наружный слой эктодермального происхождения – эпидермис; внутренний соединительнотканный слой мезодермального происхождения – кориум.
Покровы хордовых способны к образованию различных производных. У низших хордовых в их развитии участвует в основном эпидермис, у позвоночных покровы формируются чаще всего из эпидермиса и кориума.
Строение и особенности функционирования покровов тесно связаны с образом жизни, который ведет конкретная группа хордовых. В частности, принципиальные различия имеются в покровах водных и наземных позвоночных.
Покровы низших хордовых Бесчерепные
Эпидермис однослойный, кориум слабо выражен – студенистый, представлен тонким слоем бесструктурной соединительной ткани, в которой практически отсутствуют клеточные элементы и волокна. Хорошо выражена железистая функция покровов. Одноклеточные эпидермальные железы выделяют на поверхность покровов мукополисахаридную
слизевую плёнку, которая защищает покровы от повреждения частицами грунта.
Оболочники
Эпидермис оболочников выделяет на поверхность плотную оболочку – Тунику, состоящую из клетчаткоподобного вещества полисахарида – туницина, белков и небольшого количества минеральных солей. Под эпидермисом располагается кориум, представленный рыхлой соединительной тканью.
Общие особенности покровов позвоночных
Для всех позвоночных характерны многослойный эпидермис и хорошо структурированный кориум, содержащий клеточные и волокнистые элементы.
Покровы позвоночных чаще всего пигментированы. Пигменты, обеспечивающие различную окраску тела, могут располагаться как в эпидермисе, так и в клетках кориума. Имеются специализированные пигментные клетки хроматофоры, которые являются производными ганглионарной пластики. Это отростчатые, подвижные клетки.
Покровы позвоночных способны формировать различные производные при участии как эпидермиса, так и кориума. Довольно значительными особенностями обладают покровы Анамний и Амниот.
Для покровов Анамний характерно: отсутствие в покровах ороговеваний, обилие кожных желез, слизевая плёнка, уменьшающая трение и выполняющая защитную функцию, правильное сетеобразное расположение волокон кориума, повышающее эластичность (рис. 1).
Амниоты – организмы, хорошо приспособленные к наземной среде обитания, для их покровов характерно: ороговевание эпидермиса, ослабление железистой функции покровов, различные роговые образования, в формировании которых преобладает роль эпидермиса, неправильное расположение волокон кориума.
Сравнительная анатомия покровов позвоночных
Круглоротые
У современных круглоротых наружный скелет отсутствует. Роговые производные покровов чрезвычайно редки (роговые зубы воронки). Много одноклеточных желез, иногда имеются ядовитые железы. У миксин эпидермис превращен в сплошной слизистый слой – stratum mucosum.
Хрящевые рыбы
Наружный скелет представлен плакоидными чешуями. Плакоидная чешуя имеет вид шипа, выступающего наружу и имеющего основание в виде ромбовидной пластинки, расположенной в кориуме (рис. 2). Плакоидная чешуя образована в основном дентином, который формируют клетки
Дентин
шипа
Склеробласт
Основная
пластинка
Рис.
2.
Развитие плакоиднои чешуи.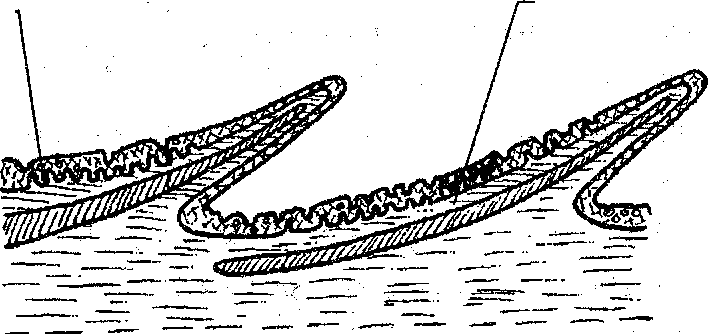
Эпидермис.
.Костная
чешуя
Рис.
3.
Разрез через кожу костистой рыбы.
Бокаловидные клетки
Зернистые
клетки
Колбовидные
клетки
Основной
слой эпидермиса
Кориум
Нервы
Сосуды
Рис.
1.
Схематичный разрез через кожу рыбы.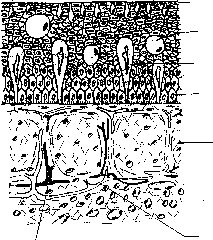
!
кориума – одонтобласты. Дентин – обызвествлённое органическое вещество, не содержащее клеток, но пронизанное каналами, в которые заходят отростки одонтобластов. Снаружи шип покрыт чрезвычайно плотным веществом – дуродентином {эмалью), который так же формируется одонтобластами. Эмаль не имеет каналов и значительно прочнее дентина.
Костные рыбы
У костных рыб встречаются три типа чешуи: ганоидная, космоидная и костная (рис. 3,4).
Ганоидная чешуя характерна для костных ганоидов (панцирная щука) и многоперых рыб. Этот тип чешуи представляет собой твердые пластинки, тесно соединяющиеся друг с другом в виде панциря. Каждая пластинка состоит из двух слоев (рис. 4). Нижний (внутренний) слой – изопедин – близок по строению к кости. Наружный – ганоин – разновидность дентина, но по прочности близок к эмали. Оба слоя образуются при участии клеток кориума, то есть мезодермального происхождения. Возможен средний слой – космин.
Космоидная чешуя – особый вариант ганоидной чешуи. Она характерна для кистеперых рыб (и вымерших двоякодышащих). В этой чешуе (рис. 4) между слоями изопедина и ганвина расположен космин – особый вариант дентина, содержащий полости с соединительной тканью и пронизанный многочисленными каналами. Все слои образуются в кориуме.
. в •
• в • * »
„ в в « »
-Ганоин
-Изопедин
Космоидная чешуя
вв<* в
-Ганоин![]()
Ш£
-Космин
в
<*
-Изопедин
(кость)
0-
— •.
«
«
* • в
Ганоидная
чешуя
■Кость
Костная чешуя
Рис. 4. Типы чешуй.
17
Костные чешуи характерны в основном для костистых рыб. Представляют собой пластинки из костного бесклеточного вещества. Расположены в основном в кориуме, частично выступают наружу и черепицеобразно налегают друг на друга. Эта чешуя возникла в эволюции двумя различными путями: во- первых, в результате редукции космина, как у современных
двоякодышащих рыб (протоптера и лепидосирена), а во-вторых,
в результате редукции ганоина, как у лучеперых рыб. Тонкую, чисто изопединовую, эласмоидную (т. е. пластинчатую) чешую костистых рыб подразделяют на две категории — циклоидную (округлую – большинство рыб) и ктеноидную (в виде гребня – окунеобразные).
Эволюция чешуи. Наиболее удачно этот вопрос решает палеонтологическая гипотеза. Согласно этой гипотезе, исходным вариантам наружного скелета позвоночных был панцирь сложного строения, покрытый конусами, подобными по строению плакоидным чешуям. В эволюции позвоночных происходила редукция отдельных компонентов исходного панциря с преобладанием других:
утрата панциря с преобладанием конусов – плакоидная чешуя;
утрата конусов и распадение сплошного панциря на отдельные пластинки – космоидная чешуя;
дальнейшая утрата косминового слоя – ганоидная чешуя;
дальнейшая утрата слоя ганоина – костные чешуи.
Земноводные
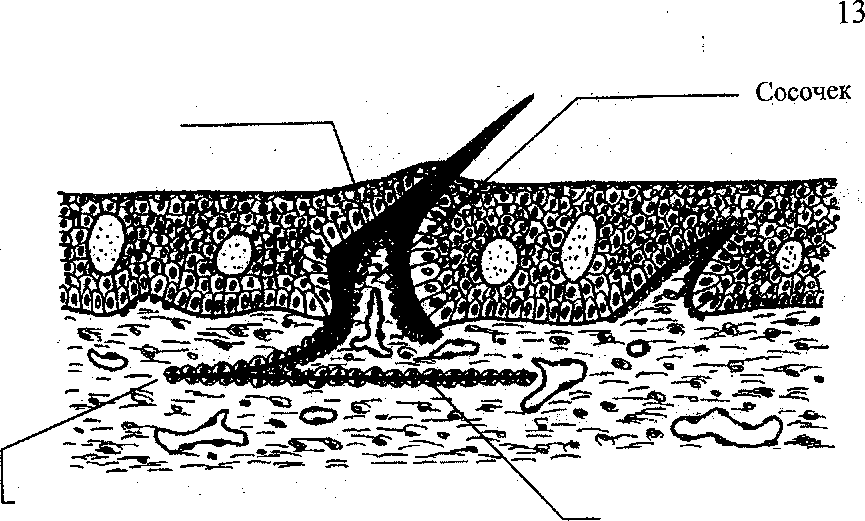
Кожа земноводных играет важную рель в газообмене, поэтому в ней нет кожных скелетных структур и имеется большое количество многоклеточных желез (у личинок железы одноклеточные). Секрет желез увлажняет кожу, обладает бактерицидным действием, может содержать ядовитые и пахучие компоненты. Волокна кориума хорошо упорядочены (рис. 5).
Рептилии
Покровы рептилий обладают всеми особенностями покровов амниот. Характерной чертой покровов пресмыкающихся является наличие роговых чешуй различной формы (бугорчатые, черепицеобразные, чешуи в виде щитков) (рис. 6). Под ними кожные окостенения - остеодермы (рис. 6).
Желез очень мало, имеются только высокоспециализированные формы желёз (бедренные поры ящериц, мускусная железа на нижней челюсти крокодила, «анальные» железы гаттерии).
Птицы
Кожа тонкая. Окостенения отсутствуют. Желез мало (копчиковая, серные железы в слуховом проходе куриных). Много роговых образований: роговые чешуи, когти, роговой клюв, перья (рис. 7).
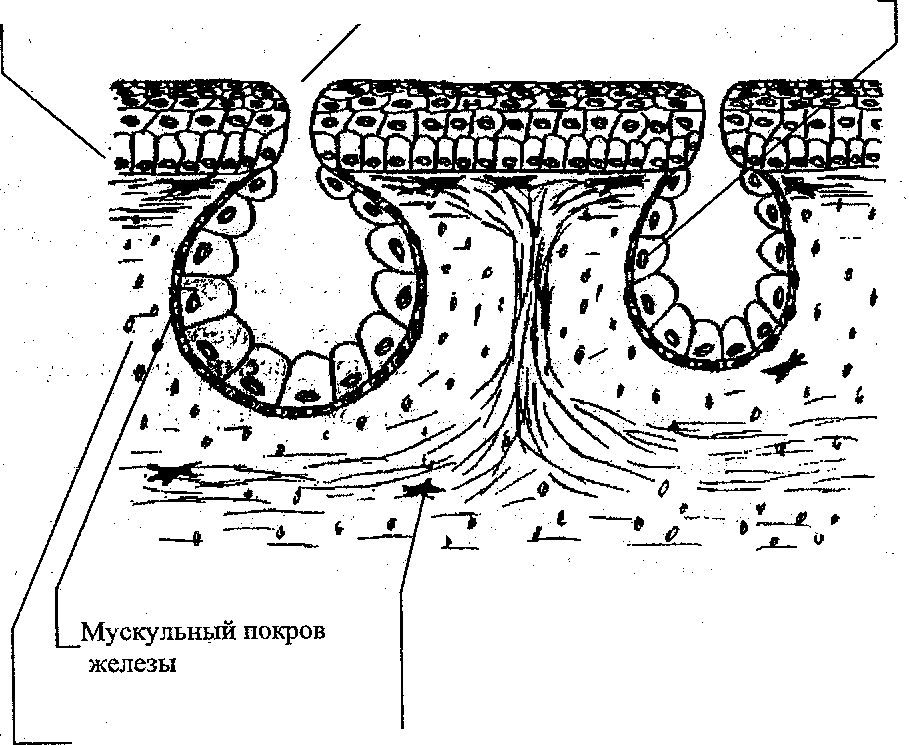
Проток
железы Железистые клетки
Кориум
Рис.
5.
Разрез через кожу амфибий.
Эпидермис
Бугорчатые чешуи
Щитки
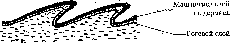
Черепицеобразные чешуи
Мальпигиев
слой
Черепицеобразные
чешуи
Рис.
6.
Схемы роговых чешуй.
Пигментные
клетки
Сраенителъная
анатомия покровов
Перья представляют собой главным образом роговые эпидермальные структуры, возникшие в процессе эволюции, как полагают, из чешуи рептилий. Они выполняют две важные функции: во-первых, покрывая все тело птицы, перья обеспечивают эффективную теплоизоляцию, способствуя поддержанию высокой температуры тела, характерной для птиц и для млекопитающих; во-вторых, крупные перья, образующие большую часть поверхности крыльев и хвоста, делают возможным полет. Различают перья трех основных типов – пуховые, контурные и нитевидные. Зрелое перо целиком состоит из сильно модифицированных и ороговевших эпителиальных клеток.



Мальпигиев
слой
эпидермиса
Роговой
слой
Сосочек
пера
•Зачаток
влагалища пера
Рис.
7. Последовательные стадии развития
пера.
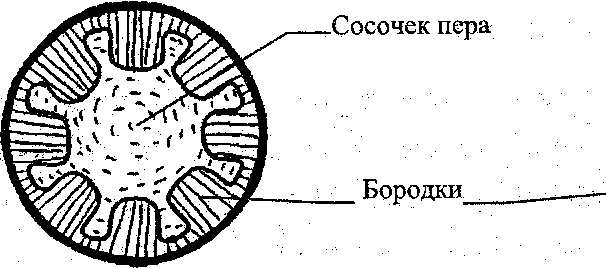
Рис.
8.
Стереограмма развивающегося контурного
пера.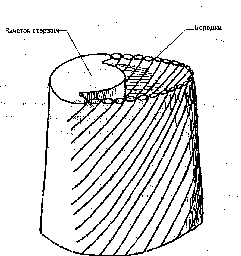
Рис.
9.
Развитие бородок эмбрионального пуха.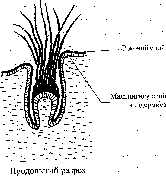
Млекопитающие
Для их покровов характерен толстый многослойный эпидермис, в котором выделяют несколько слоев: ростовой, зернистый, блестящий, роговой. Извилистая граница между кориумом и эпидермисом обеспечивает хорошую трофику эпидермиса. Хорошо развита подкожная жировая клетчатка. В коже имеется три типа желез: потовые, сальные и млечные. Потовые железы двух типов: апокриновые и эккриновые. Апокриновые потовые железы трубчатые, открываются в волосяные сумки, реагируют на адреналин, при секреции от клеток отшнуровываются капельки протоплазмы (красноватый пот бегемота, жиропот овец, «мыло» лошадей). Неповторимый набор белков обеспечивает строго индивидуальный запах. У человека эти железы деградировали, остались железы Молля на веках, серные железы в наружном слуховом проходе. Млечные железы - из этой же категории. У однопроходных они связаны с волосами, у сумчатых волосы исчезают в ходе онтогенеза, у плацентарных волосы у этих желез не закладываются.
Эккриновые потовые железы – открываются в коже независимо от волос, реагируют на ацетилхолин, секрет фильтруется через мембрану. Эти железы распространены на безволосых частях (подошвы кошек и собак). У человека они располагаются по всему телу и используются в целях терморегуляции - выделяют «дешевый» пот (без белков).
Хорошо развиты роговые производные: когти, ногти, копыта, чехлы рогов у полорогих, роговые чешуи, волосы. Онтогенез вол ос отличается от онтогенеза роговой чешуи, что свидетельствует о нешмояогичности этих образований. Развитие волос показано на рис. 10. Основной функцией волосяного покрова является теплоизоляция.