

49. Морфофункциональная характеристика ретикулярной формации
Представляет собой филогенетически более старую и относительно просто организованную нервную сеть с множеством ядерных центров. Ей отводится важная роль в поддержании бодрствующего состояния мозга, а также в механизмах формирования сложно-координированных двигательных актов (таких, как чихание, рвота и т. п.), обеспечивающих защиту организма от воздействий внешней среды, угрожающих его жизнедеятельности. Она работает в функциональном единстве с анализаторными системами и оказывает тонические влияния на ниже- и вышележащие отделы центральной нервной системы.
К числу особенностей строения ретикулярной формации следует отнести:
• глубокое и диффузное расположение ее ядер в стволе мозга; при про ведении раздражения в ретикулярной формации имеет место множе ственность переключений на ее многочисленных ядрах;
• способность проводить возбуждения в различных, как восходящих, так и нисходящих, направлениях;
• обслуживание как соматических, так и висцеральных функций;
• разнообразие групп ядер по хемоархитектонике (наличие клеточных скоплений с серотонинергическими, холинергическими, адренерги- ческими и дофаминергическими нейронами).
Ретикулярные ядра группируются в стволе мозга в три билатерально симметричных продольных столба: срединный (ядра шва), медиальный, содержащий ядра с крупными ретикулярными нейронами, и латеральный, содержащий ядра из мелких и средних по размерам нейронов (рис. 66).
Множественность и разнообразие групп ядер, входящих в состав ретикулярной формации, позволяют характеризовать ее как неспецифическую по-лисинаптическую систему. В частности, влияние ретикулярной формации на кору больших полушарий осуществляется посредством ретикуло-таламиче-ских путей через группу медиальных ядер таламуса, составляющих так называемую неспецифическую проекционную таламо-кортикальную систему. Волокна ретикуло-таламо-кортикальных путей заканчиваются во всех областях коры (так называемые неспецифические афферентные волокна коры боль шого мозга), но наибольшее их число обнаруживают в коре лобной доли.
![]()
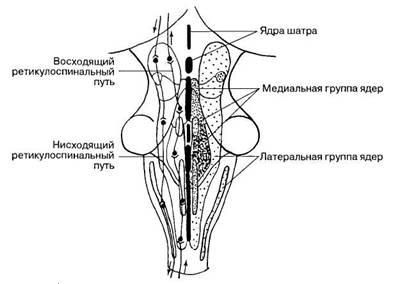
Рис. 66. Ядра ретикулярной формации в стволе мозга.
Они проводят грубую (протопатическую) чувствительность от кожного покрова и всех органов, оказывая неспецифическое активирующее воздействие на кору больших полушарий. Их называют неспецифическими афферентными волокнами в отличие от специфических, проводящих более тонкую (эпикрити-ческую) чувствительность к соматосенсорной коре, которая позволяет тонко дифференцировать локализацию и нюансы ощущений. Специфические афферентные волокна заканчиваются на вставочных нейронах IV слоя коры аксосоматическими контактами, обеспечивающими быстрые и дифференцированные ответы нейронов. Неспецифические афферентные волокна в составе восходящей активирующей системы образуют концевые ветвления во всех слоях коры мозга и формируют синаптические контакты на дендри-тах нейронов, что позволяет только модулировать возбудимость нейронов и влиять на их способность к проведению импульсов.
Активность как восходящих, так и нисходящих систем поддерживается непрерывным притоком афферентных импульсов, поступающих в ретикулярную формацию по коллатеральным волокнам от сенсорных проводящих путей. Важную роль в поддержании активности ретикулярных ядер играют гуморальные раздражители, по отношению к которым ретикулярная формация обладает высокой чувствительностью, что обеспечивает ее участие в регуляции ряда вегетативных функций.
Все двигательные ядра черепных нервов получают от ретикулярных ядер волокна, идущие в составе восходящих и двух нисходящих ретикулярных путей: мосто-спинномозгового {латеральный ретикулоспиналъный путь) и бульбо-спинномозгового {медиальный ретикулоспиналъный путь). Волокна от ретикулярных ядер заканчиваются билатерально на мотонейронах спинного мозга и симпатических преганглионарных нейронах его грудного отдела. Эти пути оказывают влияние на состояние спинномозговых мотонейронов, а также болевую чувствительность на уровне сегментарных связей.