

обмін вуглеводів
.docОБМІН
ВУГЛЕВОДІВ
9.1.
Обмін полісахаридів. Травлення
вуглеводів
Вуглеводи
займають помітне місце в раціоні людини.
Вважають, що добова потреба в них
становить 400-500 г, в п’ять разів більше,
ніж в білках. В традиційному раціоні
вуглеводи забезпечують 70 % енергетичних
потреб, хоча вважається, що на їх частку
в збалансованій дієті повинно припадати
біля 50 %енергозабезпечення.
Вуглеводи служать також джерелом
вуглецевих ланцюгів для побудови
різноманітних біомолекул.
Всі вуглеводи,
які всмоктуються з травного тракту, є
взаємозамінними, так як перетворюються
в спільні продукти - моносахариди,
переважно глюкозу, а також фруктозу,
галактозу. Можна назвати лише один
незамінний продукт вуглеводного
походження, який не синтезується в
організмі людини - аскорбінову
кислоту.
Однак велике значення для
організму має швидкість засвоєння ним
вуглеводів. Швидко засвоюються
моносахариди, особливо глюкоза. Основним
джерелом глюкози, яка швидко засвоюється,
в їжі служить сахароза, яка під впливом
ферменту сахарази розщеплюється на
глюкозу і фруктозу. Вважають, що раптове
зростання рівня глюкози в крові є
основною причиною ожиріння і діабету,
тому в раціоні таких хворих не повинно
бути легкозасвоюваної глюкози. У здорових
людей вона може становити до 100 г на
добу. '
Фруктоза засвоюється на
43 % повільніше
від глюкози. Тому в раціональному
харчуванні їй надають перевагу. Джерелом
фруктози є мед, фрукти. До порівняно
легко засвоюваних вуглеводів належить
також лактоза молока, яка гідролізує
на глюкозу і галактозу під впливом
ферменту лактази. Однак, цей фермент,
активний у дітей, у багатьох дорослих
людей втрачає активність, в результаті
чого вживання молока приводить до
порушення травлення.
Полісахариди
розщеплюються в травном
class=Section7>
у
тракті повільно, отже, не викликають
значного підйому рівня глюкози в крові.
Крім того, цінність їх в раціоні зумовлена
тим, що вони поєднуються в продуктах з
вітамінами групи В і , так званими,
харчовими волокнами. Прикладами такого
поєднання є продукти з непросіяного
борошна, квасоля, капуста та ін.
До
харчових волокон належать целюлоза і
пектини, їх багато в овочах і фруктах
(гарбуз та інші бахчеві, яблука), де вони
утворюють особливо цінні поєднання з
іншими компонентами. Пектини при
травленні розпадаються до сахарних
кислот, тому вони шкодять при деяких
захворюваннях травного тракту. Однак
для великої більшості людей вони корисні
як сорбенти токсичних речовин, які
потрапляють в травний тракт. Целюлоза
в травному тракті людини не розщеплюється,
оскільки в ньому не діють ферменти
гідролізу b-глікозидних зв’язків. Тому
довгий час присутності харчових волокон
у дієті не надавали уваги. Однак в
сучасній дієтології навіть рекомендується
збагачувати деякі продукти харчовими
волокнами, оскільки вони покращують
перистальтику кишечника, зв’язують
токсичні речовини, зокрема йони важких
металів (в тому числі і радіоактивних),
холестерин і сприяють іх виведенню з
організму.
Травлення полісахаридів
починається в ротовій порожнині під
впливом ферментів, які виділяються
слинними залозами. Великі молекули
розпадаються до менших уламків -
декстринів. Розщеплення їх продовжується
в тонкому кишечнику під впливом ферментів,
що виділяються підшлунковою залозою.
Тут декстрини руйнуються до ди- і
моносахаридів. Останні всмоктуються в
кров. Розщеплення вуглеводів в травному
тракті відбувається шляхом гідролізу.
(С6Н10О5)n
+ Н2О ® (С6Н10О5)m ® nС6Н12О6 , де m > n
Гідролази
слинних і підшлункових залоз, які
розщеплюють a-1®4
зв’язки в полісахаридах, називаються
амілазами. Вони активні в слабколужному
середовищі. Існує декілька амілаз, які
діють на молекулу полісахариду в різних
позиціях:
 a-амілаза
(1) прискорює гідроліз 1®4-глікозидних
зв’язків всередині ланцюга
b-амілаза
(2) відщеплює послідовно залишки мальтози
з нередукуючого кінця молекули.
g-амілаза
(3) відщеплює послідовно залишки глюкози
від нередукуючого кінця молекули
полісахариду.
Зв’язки 1®6 в точках
галуження полісахаридів розщеплює
фермент аміло-1,6-глюкозидаза (4).
Дисахариди
також піддаються гідролізу в травному
тракті під дією відповідних ферментів.
Наприклад, мальтоза розщеплюється під
дією ферменту мальтази на дві молекули
глюкози.
Кінцевим продуктом гідролізу
є моносахариди: переважно глюкоза, а
також фруктоза, галактоза та інші.
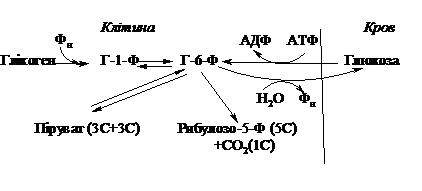
Обмін
глікогену в клітині та його регуляція
Розпад
резервного полісахариду глікогену в
клітинах печінки і м’язів суттєво
відрізняється від розпаду полісахаридів
при травленні. Він здійснюється
шляхомфосфоролізу,
тобто за участю фосфорної кислоти.
Реакцію каталізує фермент фосфорилаза,
яка відщеплює залишки глюкози з
нередукуючого кінця молекули і фосфорилює
їх з утворенням глюкозо-1-фосфату
(Г-1-Ф):
(С6Н10О5)n + Н3РО4 ® (С6Н10О5)n-1 +
Г-1-Ф
Фосфорилаза належить до класу
трансфераз. Це глікозилтрансфераза.
Вона є регуляторним ферментом і може
перебувати в двох формах: активній (а)
і неактивній (b). Алостеричним регулятором
ферменту є фосфат. Фосфорилаза b
складається з двох субодиниць. За участю
фосфату відбувається утворення активної
форми - тетрамеру. Активація ферменту
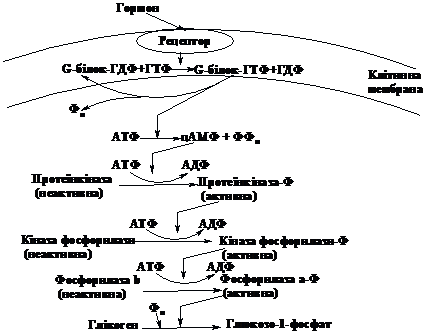
регулюється за каскадним принципом.
Фосфорилювання ферменту є лише останнім
етапом, а весь каскадний механізм
запускається дією гормонів адреналіну
(на клітини м’язів) або глюкагону (на
клітини печінки). Всередину клітини ці
гормони не проникають, а взаємодіють
із специфічними рецепторами мембрани,
стимулюючи утворення в клітині ц-АМФ.
Цей процес опосередкований мембранним
ГТФ-зв’язуючим білком (G-білком).
В мембрані цей білок існує у вигляді
комплекса з ГДФ. При взаємодії з комплексом
гормон-рецептор G-білок
обмінює молекулу ГДФ на ГТФ і в такому
вигляді стає активатором аденілатциклази.
Після активації ГТФ гідролізує до ГДФ
і G-білок
може знову виконувати свою функцію.
Ц-АМФ
активує фермент протеїнкіназу, а той,
в свою чергу, каталізує утворення
активної форми фосфорилази, яка здійснює
фосфороліз глікогену. Таким чином,
розщеплення глікогену стимулюється
гормонами і ц-АМФ відіграє в цьому
процесі роль посередника дії гормонів
в клітині.
a-амілаза
(1) прискорює гідроліз 1®4-глікозидних
зв’язків всередині ланцюга
b-амілаза
(2) відщеплює послідовно залишки мальтози
з нередукуючого кінця молекули.
g-амілаза
(3) відщеплює послідовно залишки глюкози
від нередукуючого кінця молекули
полісахариду.
Зв’язки 1®6 в точках
галуження полісахаридів розщеплює
фермент аміло-1,6-глюкозидаза (4).
Дисахариди
також піддаються гідролізу в травному
тракті під дією відповідних ферментів.
Наприклад, мальтоза розщеплюється під
дією ферменту мальтази на дві молекули
глюкози.
Кінцевим продуктом гідролізу
є моносахариди: переважно глюкоза, а
також фруктоза, галактоза та інші.
Обмін
глікогену в клітині та його регуляція
Розпад
резервного полісахариду глікогену в
клітинах печінки і м’язів суттєво
відрізняється від розпаду полісахаридів
при травленні. Він здійснюється
шляхомфосфоролізу,
тобто за участю фосфорної кислоти.
Реакцію каталізує фермент фосфорилаза,
яка відщеплює залишки глюкози з
нередукуючого кінця молекули і фосфорилює
їх з утворенням глюкозо-1-фосфату
(Г-1-Ф):
(С6Н10О5)n + Н3РО4 ® (С6Н10О5)n-1 +
Г-1-Ф
Фосфорилаза належить до класу
трансфераз. Це глікозилтрансфераза.
Вона є регуляторним ферментом і може
перебувати в двох формах: активній (а)
і неактивній (b). Алостеричним регулятором
ферменту є фосфат. Фосфорилаза b
складається з двох субодиниць. За участю
фосфату відбувається утворення активної
форми - тетрамеру. Активація ферменту
регулюється за каскадним принципом.
Фосфорилювання ферменту є лише останнім
етапом, а весь каскадний механізм
запускається дією гормонів адреналіну
(на клітини м’язів) або глюкагону (на
клітини печінки). Всередину клітини ці
гормони не проникають, а взаємодіють
із специфічними рецепторами мембрани,
стимулюючи утворення в клітині ц-АМФ.
Цей процес опосередкований мембранним
ГТФ-зв’язуючим білком (G-білком).
В мембрані цей білок існує у вигляді
комплекса з ГДФ. При взаємодії з комплексом
гормон-рецептор G-білок
обмінює молекулу ГДФ на ГТФ і в такому
вигляді стає активатором аденілатциклази.
Після активації ГТФ гідролізує до ГДФ
і G-білок
може знову виконувати свою функцію.
Ц-АМФ
активує фермент протеїнкіназу, а той,
в свою чергу, каталізує утворення
активної форми фосфорилази, яка здійснює
фосфороліз глікогену. Таким чином,
розщеплення глікогену стимулюється
гормонами і ц-АМФ відіграє в цьому
процесі роль посередника дії гормонів
в клітині.
Синтез
глікогену здійснюється з глюкозо-1-фосфату.
Процес каталізується ферментом
глікогенсинтазою (синтазами називаються
синтетази, які забезпечують реакції
синтезу без витрат АТФ). Цей фермент не
може починати синтез полісахаридного
ланцюга із об’єднання двох ланок, а
нарощує невеликий (не менше чотирьох
моносахаридних ланок) олігосахаридний
ланцюг – «затравку», приєднуючи до його
нередукуючого кінця активовані залишки
глюкози з утворенням зв’язків
a-1,4-.
Активація
глюкозо-1-фосфату полягає в утворення
уридилдифосфоглюкози:
Г-1-Ф + УТФ ®
УДФ-Г + ФФн.
 Ця
реакція каталізується
глюкозо-1-фосфат-уридилтрансферазою.
Активна форма глюкози взаємодіє з
полісахаридним ланцюгом:
(С6Н10О5)n +
УДФ-Г ® (С6Н10О5)n+1 + УДФ
Нуклеозиддифосфати
вуглеводів використовуються як активна
форма і в інших синтезах. За роботи по
вивченню механізму синтезу глікогену
аргентинський біохімік Луїс Лелуар у
1970 p. був нагороджений Нобелівською
премією. Галуження полісахаридного
ланцюга з утворенням зв’язків
a-1,6-здійснює фермент
глікозил-(4®б)-трансфераза.
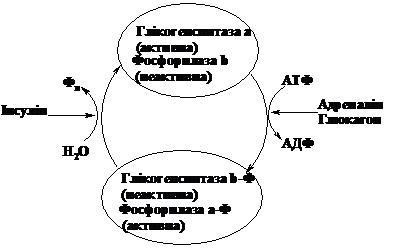
Глікогенсинтаза,
як і фосфорилаза, є регуляторним
ферментом. Спосіб її активації протилежний
до способу активації фосфорилази. В
активній формі (а) вона не фосфорильована.
Фосфорилювання під дією протеїнкінази
за рахунок АТФ приводить до утворення
неактивної форми (в).
Є відомості, що
процес активації глікогенсинтази
запускає гормон інсулін, який не проникає
в клітину, а діє на її мембрану. Підвищення
вмісту глюкози в крові стимулює виділення
інсуліну клітинами підшлункової залози.
Одним з ефектів цього гормону і є
посилення утворення глікогену, причому
найбільш швидко синтез глікогену в
печінці відбувається через 30-40 хв. після
прийому вуглеводів з їжею.
Таким чином,
розпад і синтез глікогену відбувається
узгоджено:
Ця
реакція каталізується
глюкозо-1-фосфат-уридилтрансферазою.
Активна форма глюкози взаємодіє з
полісахаридним ланцюгом:
(С6Н10О5)n +
УДФ-Г ® (С6Н10О5)n+1 + УДФ
Нуклеозиддифосфати
вуглеводів використовуються як активна
форма і в інших синтезах. За роботи по
вивченню механізму синтезу глікогену
аргентинський біохімік Луїс Лелуар у
1970 p. був нагороджений Нобелівською
премією. Галуження полісахаридного
ланцюга з утворенням зв’язків
a-1,6-здійснює фермент
глікозил-(4®б)-трансфераза.
Глікогенсинтаза,
як і фосфорилаза, є регуляторним
ферментом. Спосіб її активації протилежний
до способу активації фосфорилази. В
активній формі (а) вона не фосфорильована.
Фосфорилювання під дією протеїнкінази
за рахунок АТФ приводить до утворення
неактивної форми (в).
Є відомості, що
процес активації глікогенсинтази
запускає гормон інсулін, який не проникає
в клітину, а діє на її мембрану. Підвищення
вмісту глюкози в крові стимулює виділення
інсуліну клітинами підшлункової залози.
Одним з ефектів цього гормону і є
посилення утворення глікогену, причому
найбільш швидко синтез глікогену в
печінці відбувається через 30-40 хв. після
прийому вуглеводів з їжею.
Таким чином,
розпад і синтез глікогену відбувається
узгоджено:
 При
виконанні роботи стимулюється розпад
глікогену і пригнічується його синтез.
В стресовій ситуації (необхідність
боротьби, втечі, розумового напруження)
обмін глікогену залежить від адреналіну,
який називають “бойовим, стрімким”
гормоном. Розпад глікогену під його
впливом забезпечує енергетичними
ресурсами швидкі фізіологічні реакції
організму. Часто з метою енергозабезпечення
м’язової роботи вживають легкозасвоювані
вуглеводи (наприклад, спортсмени перед
змаганнями), Однак зростання рівня
глюкози в крові через 15-20 хв. після цього,
навпаки, сприяє її посиленій утилізації
в глікоген під впливом інсуліну і не
викликає бажаного ефекту.
9.2.
Шляхи окиснення моносахаридів
Глюкоза,
яка попадає в клітину ззовні, вступає
в перетворення лише в активній
фосфорильованій формі. Фосфорилювання
відбувається під впливом ферментів
гексокінази або глюкокінази і джерелом
фосфату служить АТФ.
При
виконанні роботи стимулюється розпад
глікогену і пригнічується його синтез.
В стресовій ситуації (необхідність
боротьби, втечі, розумового напруження)
обмін глікогену залежить від адреналіну,
який називають “бойовим, стрімким”
гормоном. Розпад глікогену під його
впливом забезпечує енергетичними
ресурсами швидкі фізіологічні реакції
організму. Часто з метою енергозабезпечення
м’язової роботи вживають легкозасвоювані
вуглеводи (наприклад, спортсмени перед
змаганнями), Однак зростання рівня
глюкози в крові через 15-20 хв. після цього,
навпаки, сприяє її посиленій утилізації
в глікоген під впливом інсуліну і не
викликає бажаного ефекту.
9.2.
Шляхи окиснення моносахаридів
Глюкоза,
яка попадає в клітину ззовні, вступає
в перетворення лише в активній
фосфорильованій формі. Фосфорилювання
відбувається під впливом ферментів
гексокінази або глюкокінази і джерелом
фосфату служить АТФ.
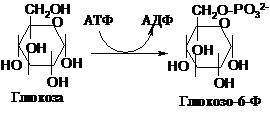 Ця
реакція протікає необоротно. Вона є
“пусковою” для різних процесів
перетворення гексоз. Глюкокіназа діє
специфічно на глюкозу. У хворих на
цукровий діабет її активність понижена,
що є одним з факторів, що протидіють
засвоєнню глюкози клітинами. Гексокіназа
фосфорилює і інші гексози. Глюкозо-6-фосфат
(Г-6-Ф) є центральним метаболітом в
перетворенні гексоз в клітині:
Ця
реакція протікає необоротно. Вона є
“пусковою” для різних процесів
перетворення гексоз. Глюкокіназа діє
специфічно на глюкозу. У хворих на
цукровий діабет її активність понижена,
що є одним з факторів, що протидіють
засвоєнню глюкози клітинами. Гексокіназа
фосфорилює і інші гексози. Глюкозо-6-фосфат
(Г-6-Ф) є центральним метаболітом в
перетворенні гексоз в клітині:
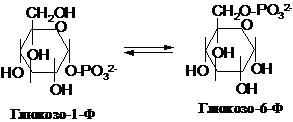
 Продукт
фосфоролізу глікогену – глюкозо-1-фосфат
(Г-1-Ф) - піддається ізомеризації за
допомогою фосфоглюкомутази:
Продукт
фосфоролізу глікогену – глюкозо-1-фосфат
(Г-1-Ф) - піддається ізомеризації за
допомогою фосфоглюкомутази:
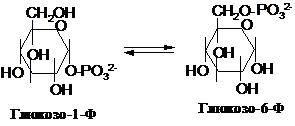 Існує
два основних способи розпаду
глюкозо-6-фосфату: дихотомічний і
апотомічний. При дихотомічному способі
молекула глюкози руйнується на дві
рівні частини - тріози, які піддаються
дальшому окисненню. При апотомічному
глюкоза, окиснюючись, розпадається на
два нерівних фрагменти: одновуглецевий
(СО2) і п’ятивуглецевий (пентозу).
Більша
частина глюкози в організмі розпадається
дихотомічним способом, з виділенням
АТФ для забезпечення енергетичних
потреб.
Дихотомічнерозщеплення
глюкози більше
відоме під назвою “гліколіз” (від
“лізіс” - розщеплення). Це один з
еволюційно найдавніших катаболічних
шляхів. Він складається з десяти
послідовних реакцій, які уожна умовно
поділити на два етапи. На першому етапі
відбувається активація молекули за
рахунок АТФ і розщеплення активованого
продукту на дві фосфорильовані тріози.
На другому етапі утворені тріози
окиснюються з відновленням НАД+. Проміжні
продукти окиснення є супермакроергами
і забезпечують синтез АТФ. Кінцевим
продуктом дихотомічного розпаду є
піруват (ПК). Отже, перший етап є
підготовчим, а другий реалізує можливості
глюкози для одержання енергії і відновних
еквівалентів.
Дихотомічний шлях
починається з ізомеризації глюкозо-6-фосфату
у фруктозо-6-фосфат (Фр-б-Ф) за участю
фосфоглюкоізомерази:
Існує
два основних способи розпаду
глюкозо-6-фосфату: дихотомічний і
апотомічний. При дихотомічному способі
молекула глюкози руйнується на дві
рівні частини - тріози, які піддаються
дальшому окисненню. При апотомічному
глюкоза, окиснюючись, розпадається на
два нерівних фрагменти: одновуглецевий
(СО2) і п’ятивуглецевий (пентозу).
Більша
частина глюкози в організмі розпадається
дихотомічним способом, з виділенням
АТФ для забезпечення енергетичних
потреб.
Дихотомічнерозщеплення
глюкози більше
відоме під назвою “гліколіз” (від
“лізіс” - розщеплення). Це один з
еволюційно найдавніших катаболічних
шляхів. Він складається з десяти
послідовних реакцій, які уожна умовно
поділити на два етапи. На першому етапі
відбувається активація молекули за
рахунок АТФ і розщеплення активованого
продукту на дві фосфорильовані тріози.
На другому етапі утворені тріози
окиснюються з відновленням НАД+. Проміжні
продукти окиснення є супермакроергами
і забезпечують синтез АТФ. Кінцевим
продуктом дихотомічного розпаду є
піруват (ПК). Отже, перший етап є
підготовчим, а другий реалізує можливості
глюкози для одержання енергії і відновних
еквівалентів.
Дихотомічний шлях
починається з ізомеризації глюкозо-6-фосфату
у фруктозо-6-фосфат (Фр-б-Ф) за участю
фосфоглюкоізомерази:
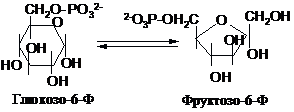 Наступна
реакція є другою “пусковою” на шляху
перетворень -це фосфорилювання
фруктозо-6-фосфату за рахунок АТФ:
Наступна
реакція є другою “пусковою” на шляху
перетворень -це фосфорилювання
фруктозо-6-фосфату за рахунок АТФ:

В
клітині ця реакція необоротна. Утворений
фруктозо-1,6-дифосфат (Фр-1,6-диФ) є майже
симетричною молекулою з двома однойменно
зарядженими фосфатними групіровками,
тому він легко розщеплюється за участю
ферменту альдолази на дві тріози:
3-фосфогліцериновий альдегід (3-ФГА) і
диоксиацетонфосфат (ДОАФ):
 Ці
тріози легко ізомеризуються
тріозофосфатізомеразою:
Ці
тріози легко ізомеризуються
тріозофосфатізомеразою:
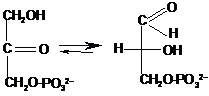 На
цьому закінчується перший етап
дихотомічного розпаду. Витрати енергії
на ньому становлять 1 АТФ на 1 молекулу
глюкозофосфату, або 2 АТФ на 1 молекулу
глюкози.
В перетворення другого етапу
вступає лише один з двох продуктів -
3-фосфогліцериновий альдегід. Тому
рівновага в реакції ізомеризації тріоз
зміщена в бік утворення альдегіду. Отже,
одна молекула глюкози дає дві молекули
3-фосфогліцеринового альдегіду для
наступних перетворень.
Другий етап
починається з реакції окиснення альдегіду
в кислоту, причому за участю фосфату
альдегідна група окиснюється не просто
в карбоксильну, а в карбоксилфосфатну
(змішаний ангідрид двох кислот). Утворений
продукт - це 1,3-дифосфогліцеринова
кислота (1,3-ДФГ). Акцептором гідрогену
від окиснюваної групи є НАД+. Ця складна
реакція каталізується ферментом
гліцеральдегідфос-фатдегідрогеназою:
На
цьому закінчується перший етап
дихотомічного розпаду. Витрати енергії
на ньому становлять 1 АТФ на 1 молекулу
глюкозофосфату, або 2 АТФ на 1 молекулу
глюкози.
В перетворення другого етапу
вступає лише один з двох продуктів -
3-фосфогліцериновий альдегід. Тому
рівновага в реакції ізомеризації тріоз
зміщена в бік утворення альдегіду. Отже,
одна молекула глюкози дає дві молекули
3-фосфогліцеринового альдегіду для
наступних перетворень.
Другий етап
починається з реакції окиснення альдегіду
в кислоту, причому за участю фосфату
альдегідна група окиснюється не просто
в карбоксильну, а в карбоксилфосфатну
(змішаний ангідрид двох кислот). Утворений
продукт - це 1,3-дифосфогліцеринова
кислота (1,3-ДФГ). Акцептором гідрогену
від окиснюваної групи є НАД+. Ця складна
реакція каталізується ферментом
гліцеральдегідфос-фатдегідрогеназою:
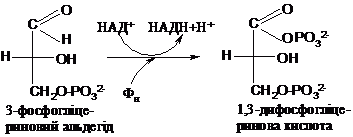 Карбоксилфосфатна
група є супермакроергічною, DG0 її
гідролізу становить - 49,3 кДж. Ця енергія
використовується для синтезу АТФ в
спряженій реакції, яку каталізує
фосфогліцераткіназа:
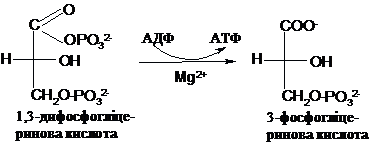
Карбоксилфосфатна
група є супермакроергічною, DG0 її
гідролізу становить - 49,3 кДж. Ця енергія
використовується для синтезу АТФ в
спряженій реакції, яку каталізує
фосфогліцераткіназа:
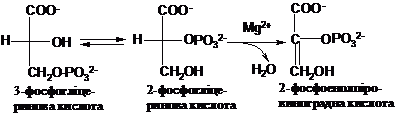
 Утворений
продукт, З-фосфогліцеринова кислота
(З-ФГК) не є макроергічною речовиною,
однак внаслідок ізомеризації в
2-фосфогліцеринову кислоту (2-ФГК) під
впливом фосфогліцератмутази і наступної
її дегідратації під впливом енолази,
вона перетворюється в супермакроергічну
сполуку - 2-фосфоенолпіровиноградну
кислоту (2-ФЕП):
Утворений
продукт, З-фосфогліцеринова кислота
(З-ФГК) не є макроергічною речовиною,
однак внаслідок ізомеризації в
2-фосфогліцеринову кислоту (2-ФГК) під
впливом фосфогліцератмутази і наступної
її дегідратації під впливом енолази,
вона перетворюється в супермакроергічну
сполуку - 2-фосфоенолпіровиноградну
кислоту (2-ФЕП):
 Енергія
її гідролізу (DG0 = -61,9 кДж) частково
використовується для синтезу АТФ в
спряженій реакції, а частково - для
незворотної ізомеризації енолпіровиноградної
кислоти (ЕПК) в кето-форму - піровиноградну
кислоту (ПК), яка переважає в умовах
клітини:
Енергія
її гідролізу (DG0 = -61,9 кДж) частково
використовується для синтезу АТФ в
спряженій реакції, а частково - для
незворотної ізомеризації енолпіровиноградної
кислоти (ЕПК) в кето-форму - піровиноградну
кислоту (ПК), яка переважає в умовах
клітини:
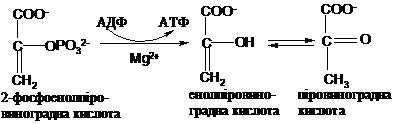 Фермент
піруваткіназа, який каталізує цей
комплекс перетворень, належить до
важливих регуляторних ферментів.
На
другій стадії дихотомічного розщеплення
в реакціях субстратного фосфорилювання
утворюється 4 молекули АТФ і 2 молекули
НАДН в розрахунку на одну молекулу
глюкози. Вся послідовність реакцій
дихотомічного розщеплення глюкози
виглядає так:
Фермент
піруваткіназа, який каталізує цей
комплекс перетворень, належить до
важливих регуляторних ферментів.
На
другій стадії дихотомічного розщеплення
в реакціях субстратного фосфорилювання
утворюється 4 молекули АТФ і 2 молекули
НАДН в розрахунку на одну молекулу
глюкози. Вся послідовність реакцій
дихотомічного розщеплення глюкози
виглядає так:
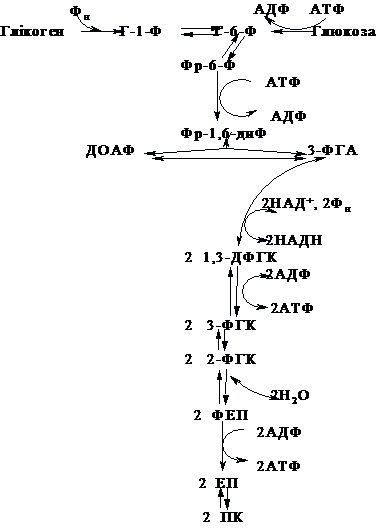 Енергетичний
баланс дихотомічного шляху, який включає
витрати на активацію молекули і вилучення
енергії з активованих продуктів, складає
+ЗАТФ для молекули ендогенного
глюкозо-1-фосфату та +2АТФ для екзогенної
молекули глюкози, на активацію якої
затрачено додатково 1 молекулу АТФ.
Необхідно врахувати також утворення
НАДН, який є джерелом відновних
еквівалентів для анаболічних реакцій,
або може стати джерелом різних форм
енергії при аеробному окиснен
class=Section8>
ні
в дихальному ланцюгу мітохондрій. З
врахуванням енергетичного балансу
сумарне рівняння дихотомічного розпаду
для глюкозофосфату можна записати
так:
Г-6-Ф + З AДФ + 2 Фн + 2 НАД+ ® 2 ПК + З АТФ
+ 2 НАДН + 2Н+,
а для глюкози:
Г + 2 AДФ + 2
Фн + 2 НАД+ ® 2 ПК + 2 АТФ + 2 НАДН +
2Н+.
Інтенсивність дихотомічного
розщеплення глюкози може змінюватись
в сотні разів завдяки алостеричній
регуляції ферментів. Фосфофруктокіназа
активується АДФ, АМФ, Фн , тобто субстратами
дихотомічного шляху, і інгібується його
продуктами: АТФ і лимонною кислотою
(один з метаболітів дальшого перетворення
піровиноградної кислоти). Аналогічно,
піруваткіназа інгібується АТФ і
ацетил-КоА, які накопичуються при
дальшому окисленні піровиноградної
кислоти.
Крім глюкози дихотомічним
шляхом можуть окиснюватись і інші
моносахариди. Галактоза за участю УТФ
перетворюється в глюкозо-1-фосфат, а
маноза і фруктоза, які фосфорилюються
під дією гексокінази, перетворюються
у фруктозо-6-фосфат (маноза попередньо
ізомеризується), які і підлягають дальшим
перетворенням, спільним з глюкозою.
Апотомічне
розщеплення глюкози.
Його поширена назва - пентозофосфатний
шлях (за назвою метаболітів). В еритроцитах,
клітинах печінки, жирової тканини та
деяких інших реакції пентозофосфатного
шляху забезпечують важливі потреби.
При окисненні глюкози акцептором
гідрогену служить на цьому шляху НАДФ+.
Утворений НАДФН є поставником гідрогену
для синтезів жирних кислот, нуклеотидів,
а також окиснення токсичних речовин в
мікросомах клітин печінки. Він захищає
від окиснення залізо гемоглобіну.
Пентозофосфатний
шлях є також джерелом пентоз для синтезів
нуклеотидів і для фотосинтезу у
рослин.
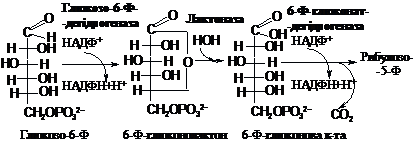
Реакції пентозофосфатного
шляху можна поділити на два етапи: перший
– етап окиснення і декарбоксилювання
і другий – етап міжмолекулярних
перебудов, в результаті яких реконструюється
молекула глюкози:
І етап:
Енергетичний
баланс дихотомічного шляху, який включає
витрати на активацію молекули і вилучення
енергії з активованих продуктів, складає
+ЗАТФ для молекули ендогенного
глюкозо-1-фосфату та +2АТФ для екзогенної
молекули глюкози, на активацію якої
затрачено додатково 1 молекулу АТФ.
Необхідно врахувати також утворення
НАДН, який є джерелом відновних
еквівалентів для анаболічних реакцій,
або може стати джерелом різних форм
енергії при аеробному окиснен
class=Section8>
ні
в дихальному ланцюгу мітохондрій. З
врахуванням енергетичного балансу
сумарне рівняння дихотомічного розпаду
для глюкозофосфату можна записати
так:
Г-6-Ф + З AДФ + 2 Фн + 2 НАД+ ® 2 ПК + З АТФ
+ 2 НАДН + 2Н+,
а для глюкози:
Г + 2 AДФ + 2
Фн + 2 НАД+ ® 2 ПК + 2 АТФ + 2 НАДН +
2Н+.
Інтенсивність дихотомічного
розщеплення глюкози може змінюватись
в сотні разів завдяки алостеричній
регуляції ферментів. Фосфофруктокіназа
активується АДФ, АМФ, Фн , тобто субстратами
дихотомічного шляху, і інгібується його
продуктами: АТФ і лимонною кислотою
(один з метаболітів дальшого перетворення
піровиноградної кислоти). Аналогічно,
піруваткіназа інгібується АТФ і
ацетил-КоА, які накопичуються при
дальшому окисленні піровиноградної
кислоти.
Крім глюкози дихотомічним
шляхом можуть окиснюватись і інші
моносахариди. Галактоза за участю УТФ
перетворюється в глюкозо-1-фосфат, а
маноза і фруктоза, які фосфорилюються
під дією гексокінази, перетворюються
у фруктозо-6-фосфат (маноза попередньо
ізомеризується), які і підлягають дальшим
перетворенням, спільним з глюкозою.
Апотомічне
розщеплення глюкози.
Його поширена назва - пентозофосфатний
шлях (за назвою метаболітів). В еритроцитах,
клітинах печінки, жирової тканини та
деяких інших реакції пентозофосфатного
шляху забезпечують важливі потреби.
При окисненні глюкози акцептором
гідрогену служить на цьому шляху НАДФ+.
Утворений НАДФН є поставником гідрогену
для синтезів жирних кислот, нуклеотидів,
а також окиснення токсичних речовин в
мікросомах клітин печінки. Він захищає
від окиснення залізо гемоглобіну.
Пентозофосфатний
шлях є також джерелом пентоз для синтезів
нуклеотидів і для фотосинтезу у
рослин.
Реакції пентозофосфатного
шляху можна поділити на два етапи: перший
– етап окиснення і декарбоксилювання
і другий – етап міжмолекулярних
перебудов, в результаті яких реконструюється
молекула глюкози:
І етап:
 ІІ
етап:
ІІ
етап:
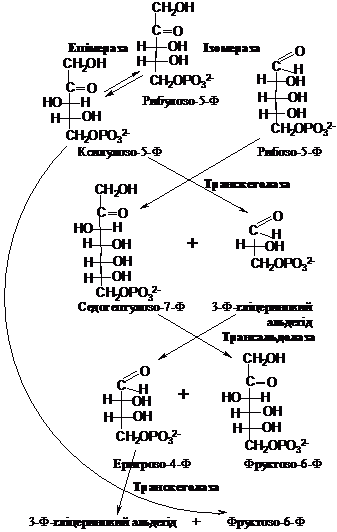
Сумарне
рівняння його можна записати так:
С6Н12О6
+ 2НАДФ+ + Н2О ® С5Н10О5 + СО2 + 2НАДФН + 2Н+
В
кінцевому рахунку реакції пентозофосфатного
шляху знову приводять до утворення
глюкози, причому з 6 молекул глюкози
одна окиснюється повністю до СО2, а п’ять
- ресинтезується:
6С6Н12О6 + 12НАДФ+ + 6Н2О
® 5С6Н12О6 + 6СО2 + 12НАДФН + 12Н+
Перетворення
піровиноградної кислоти
Піровиноградна
кислота може, залежно від умов і виду
організму, перетворюватись трьома
різними шляхами. Основним з них є аеробне
окиснення, яке
відбувається в присутності кисню. Під
впливом ферментів піруватдегідрогеназного
комплексу, до складу яких входять
коферменти тіамінпірофосфат (ТПФ), ФАД,
НАД, коензим A (KoASH), ліпоєва кислота (ЛК)
відбувається окисне декарбоксилювання
піровиноградної кислоти в ацетил-КоА
(активовану форму оцтової кислоти). Цей
процес включає декарбоксилювання
пірувату за участю ТПФ, окиснення
утвореного оцтового альдегіду, спряжене
з відновленням ЛК, і активацію окисненого
продукту коензимом А:
 Відновлена
форма ЛК забезпечує відновлення ФАД в
складі ферменту, а ФАН2, в свою чергу
відновлює НАД:
ЛК(SH)2 + ФАД ® ЛКS2 +
ФАДН2,
ФАДН2 + НАД+ ® ФАД + НАДН + Н+
Сумарне
рівняння аеробного окиснення
пірувату:
СН3СОСОО- + КоАSH + НАД+ ®
СН3СОSКоА + СО2 + НАДН + Н+
Ацетил-КоА є
спільним продуктом катаболічних шляхів.
Наступне його окиснення відбувається
в мітохондріях в цик
class=Section9>
лі
трикарбонових кислот. Детально цей цикл
буде розглянутий після вивчення
катаболізму ліпідів і амінокислот.
Зараз лише відзначимо, що повне окислення
в ньому ацетилу до двох молекул СО2 дає
максимальний енергетичний ефект 12
макроергічних фосфатних груп.
Для
обчислення повного енергетичного
балансу аеробного окиснення глюкози
приймемо, що перенесення двох атомів
гідрогену від НАДН до кисню по дихальному
шляху мітохондрій супроводжується
утворенням максимально трьох молекул
АТФ шляхом окисного фосфорилювання
(Р/О=3). Таким чином, енергетичний баланс
аеробного окислення глюкози становить
:
- 2 АТФ (активація глюкози)
+ 4 АТФ
(субстратне фосфорилювання в дихальному
ланцюгу)
+ 6 АТФ (2х3 – за рахунок 2НАДН,
відновленого в дихотомічному шляху)
+
6 АТФ (2х3 – за рахунок 2НАДН, відновленого
в піруватдегідрогеназній реакції)
+
24 АТФ (в циклі трикарбонових кислот)
Отже
сумарний ефект становить 38 молекул АТФ
(або макроергічних фосфатних груп), які
утворюються при повному окисненні
однієї молекули глюкози.
В анаеробних
умовах піруват
відновлюється до лактату (молочної
кислоти) за рахунок НАДН. Ця оборотна
реа
class=Section10>
кція
каталізується
class=Section11>
лактатдегідрогеназою:
Відновлена
форма ЛК забезпечує відновлення ФАД в
складі ферменту, а ФАН2, в свою чергу
відновлює НАД:
ЛК(SH)2 + ФАД ® ЛКS2 +
ФАДН2,
ФАДН2 + НАД+ ® ФАД + НАДН + Н+
Сумарне
рівняння аеробного окиснення
пірувату:
СН3СОСОО- + КоАSH + НАД+ ®
СН3СОSКоА + СО2 + НАДН + Н+
Ацетил-КоА є
спільним продуктом катаболічних шляхів.
Наступне його окиснення відбувається
в мітохондріях в цик
class=Section9>
лі
трикарбонових кислот. Детально цей цикл
буде розглянутий після вивчення
катаболізму ліпідів і амінокислот.
Зараз лише відзначимо, що повне окислення
в ньому ацетилу до двох молекул СО2 дає
максимальний енергетичний ефект 12
макроергічних фосфатних груп.
Для
обчислення повного енергетичного
балансу аеробного окиснення глюкози
приймемо, що перенесення двох атомів
гідрогену від НАДН до кисню по дихальному
шляху мітохондрій супроводжується
утворенням максимально трьох молекул
АТФ шляхом окисного фосфорилювання
(Р/О=3). Таким чином, енергетичний баланс
аеробного окислення глюкози становить
:
- 2 АТФ (активація глюкози)
+ 4 АТФ
(субстратне фосфорилювання в дихальному
ланцюгу)
+ 6 АТФ (2х3 – за рахунок 2НАДН,
відновленого в дихотомічному шляху)
+
6 АТФ (2х3 – за рахунок 2НАДН, відновленого
в піруватдегідрогеназній реакції)
+
24 АТФ (в циклі трикарбонових кислот)
Отже
сумарний ефект становить 38 молекул АТФ
(або макроергічних фосфатних груп), які
утворюються при повному окисненні
однієї молекули глюкози.
В анаеробних
умовах піруват
відновлюється до лактату (молочної
кислоти) за рахунок НАДН. Ця оборотна
реа
class=Section10>
кція
каталізується
class=Section11>
лактатдегідрогеназою:
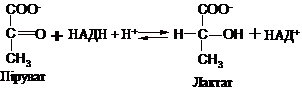 Джерелом
НАДН для відновлення піровиноградної
кислоти служить реакція окиснення
3-фосфогліцеринового альдегіду. Таким
чином, в анаеробному розщепленні глюкози
НАДН використовується для відновленення
не кисню, а пірувату. Відмінність у
використанні НАДН в аеробних і анаеробних
умовах показана на схемі:
Джерелом
НАДН для відновлення піровиноградної
кислоти служить реакція окиснення
3-фосфогліцеринового альдегіду. Таким
чином, в анаеробному розщепленні глюкози
НАДН використовується для відновленення
не кисню, а пірувату. Відмінність у
використанні НАДН в аеробних і анаеробних
умовах показана на схемі:
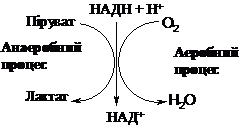
Процес
анаеробного окиснення глюкози до
молочної кислоти називаєтьсягліколізом (від
“лізіс” – розщеплення), а процес
анаеробного окиснення глікогену до
молочної кислоти - глікогенолізом. Часто
термін “гліколіз” застосовують до
дихотомічного шляху розщеплення глюкози
незалежно від умов його протікання.
Енергетичний ефект гліколізу становить
2 АТФ на одну молекулу глюкози, енергетичний
ефект глікогенолізу - З АТФ на один
залишок глюкози. Сумарне рівняння
гліколізу:
С6Н12О6 + 2АДФ + 2 Фн ® 2С3Н6О3 +
2 АТФ + 2Н2О.
Відновлення піровиноградної
кислоти до молочної, яке здійснюється
м
class=Section12>
олочнокислими
бактеріями в анаеробних умовах,
називається молочнокислим
бродін
class=Section13>
ням
Незважаючи
на невеликий,
class=Section14>
порівняно
з аеробним процесом, енергетичний ефект,
гліколіз відіграє важливе значення в
енергозабезпеченні фізичних процесів.
При потужній роботі в м’язах швидко
створюються умови дефіциту кисню і
гліколіз стає основним енергетичним
ресурсом для її забезпечення. Так у
спортсменів-бігунів на дистанції 100 м
за 10-12 с в організмі створюється дефіцит
кисню 90-95 % від
кисневого запиту і енергозабезпечення
здійснюється переважно гліколітично.
Ферменти гліколізу високоактивні у
птахів, які для захисту використовують
швидкий, панічний зліт (куріпка, фазан).
У людей і тварин, які використовують
довготривалу фізичну роботу (наприклад,
спортсмени-марафонці) виробляються
пристосування для більш ефективного
протікання аеробних процесів (більша
кількість мітохондрій, дихальних
білків).
В нормальних клітинах в
присутності кисню має перевагу аеробне
окиснення глюкози (ефект Пастера). У
ракових клітинах гліколіз не пригнічується
і в аеробних умовах.
Недоліком гліколізу,
крім малої потужності, є підкислення
середовища внаслідок утворення молочної
кислоти. Її рівень в крові може
зростати
class=Section15>
в
10 разів і більше.
Реакція утворення
молочної кислоти зворотня і в кисневих
умовах рівновага зміщується в бік
утворення пірувату з відновленням
НАД.
В клітинах дріжджів та інших
мікроорганізмів в анаеробних умовах
відбувається перетворення піровиноградної
кислоти, як
class=Section16>
е
називається спиртовим
бродінням. Фермент
піруватдекарбоксилаза, який міститься
в цих клітинах, здійснює декарбоксилювання
піровиноградної кислоти з утворенням
оцтового альдегіду: До складу
піруватдекарбоксилази входить
тіамінпірофосфат. Оцтовий альдегід
відновлюється ферментом алкогольдегідрогеназою
за участю НАДН до етанолу:
 Реакція
відновлення оцтового альдегіду зворотня.
В клітинах печінки людини алкогольдегідрогеназа
забезпечує окиснення етанолу, який в
незначній кількості утворюється в
організмі (0,001-0,01 г/л), або може попадати
ззовні. Це окиснення відбувається в дві
стадії:
Реакція
відновлення оцтового альдегіду зворотня.
В клітинах печінки людини алкогольдегідрогеназа
забезпечує окиснення етанолу, який в
незначній кількості утворюється в
організмі (0,001-0,01 г/л), або може попадати
ззовні. Це окиснення відбувається в дві
стадії:
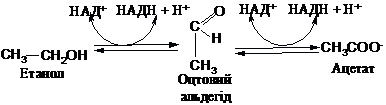 Кінцевий
продукт - оцтова кислота (ацетил-КоА) -
є природним метаболітом. Його накопичення
за рахунок окиснення етанолу, так як і
накопичення НАДН, веде до посилення
синтетичних процесів і, в кінцевому
рахунку, до жирового переродження
печінки. Окиснення надлишкового НАДН
потребує витрат кисню, внаслідок чого
виникає тканева гіпоксія. При гострому
отруєнні алкоголем особливо небезпечним
є проміжний продукт окиснення - оцтовий
альдегід - надзвичайно токсична речовина.
Один з методів лікування алкоголізму
грунтується на введенні препарату, який
блокує окислення оцтового альдегіду.
Хворий, якому введений препарат, знає,
що для нього ковток алкоголю рівнозначний
ковтку отрути.
Узагальнена схема
можливих перетворень вуглецевого
ланцюга глюкози по дихотомічному шляху
представлена на схемі:
Кінцевий
продукт - оцтова кислота (ацетил-КоА) -
є природним метаболітом. Його накопичення
за рахунок окиснення етанолу, так як і
накопичення НАДН, веде до посилення
синтетичних процесів і, в кінцевому
рахунку, до жирового переродження
печінки. Окиснення надлишкового НАДН
потребує витрат кисню, внаслідок чого
виникає тканева гіпоксія. При гострому
отруєнні алкоголем особливо небезпечним
є проміжний продукт окиснення - оцтовий
альдегід - надзвичайно токсична речовина.
Один з методів лікування алкоголізму
грунтується на введенні препарату, який
блокує окислення оцтового альдегіду.
Хворий, якому введений препарат, знає,
що для нього ковток алкоголю рівнозначний
ковтку отрути.
Узагальнена схема
можливих перетворень вуглецевого
ланцюга глюкози по дихотомічному шляху
представлена на схемі:
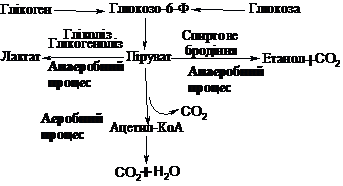 9.3.
Первинний і вторинний синтез
моносахаридів
Синтез
глюкози з низьмолекулярних
речо
class=Section17>
вин
здійснюється як рослинними, так і
тваринними клітинами.
Глюкоза може
утворюватись з органічних молекул,
що
class=Section18>
містять
три атоми карбону, зокрема кінцевих
продуктів дихотомічного розщеплення.
Синтез глюкози з піровиноградної або
молочної кислот називається глюкогенезом(від
"генезис" - народження). Відповідно
утворення глікогену цим шляхом
називається глікогенезом.
Синтез глюкози з інших речовин, які
можуть перетворюватись на піруват або
інші метаболіти глюкогенезу,
називаєтьсяглюконеогенезом.
Синтез глюкози з ацетил-КоА не може
відбуватись, так як піруватдегідрогеназна
реакція практично необоротна. Однак
для глюкогенезу може використовуватись
щавелевооцтова кислота (оксалоацетат)
– один із метаболітів циклу трикарбонових
кислот, в якому перетворюється ацетил-КоА.
Таким чином атоми карбону багатьох
метаболітів безпосередньо або через
спільні проміжні шляхи можуть бути
використані для синтезу глюкози.
Більшість
реакцій гліколізу, як було показано
вище, є оберненими. При глюкогенезі вони
ідуть у оберненому напрямі. Однак реакції
гліколізу, які є незворотними, оскільки
супроводжуються великими змінами
вільної енергії, при глюкогенезі
заміняються іншими, обхідними. Таких
обхідних реакцій при глюкогенезі є три.
Перша – це синтез фосфоенолпірувату
із пірувату. Вона відбувається з
утворенням проміжного продукту –
оксалоацетату шляхом карбоксилювання
пірувату та його наступного фосфорилювання
і декарбоксилювання з утворенням
фосфоенолпірувату:
9.3.
Первинний і вторинний синтез
моносахаридів
Синтез
глюкози з низьмолекулярних
речо
class=Section17>
вин
здійснюється як рослинними, так і
тваринними клітинами.
Глюкоза може
утворюватись з органічних молекул,
що
class=Section18>
містять
три атоми карбону, зокрема кінцевих
продуктів дихотомічного розщеплення.
Синтез глюкози з піровиноградної або
молочної кислот називається глюкогенезом(від
"генезис" - народження). Відповідно
утворення глікогену цим шляхом
називається глікогенезом.
Синтез глюкози з інших речовин, які
можуть перетворюватись на піруват або
інші метаболіти глюкогенезу,
називаєтьсяглюконеогенезом.
Синтез глюкози з ацетил-КоА не може
відбуватись, так як піруватдегідрогеназна
реакція практично необоротна. Однак
для глюкогенезу може використовуватись
щавелевооцтова кислота (оксалоацетат)
– один із метаболітів циклу трикарбонових
кислот, в якому перетворюється ацетил-КоА.
Таким чином атоми карбону багатьох
метаболітів безпосередньо або через
спільні проміжні шляхи можуть бути
використані для синтезу глюкози.
Більшість
реакцій гліколізу, як було показано
вище, є оберненими. При глюкогенезі вони
ідуть у оберненому напрямі. Однак реакції
гліколізу, які є незворотними, оскільки
супроводжуються великими змінами
вільної енергії, при глюкогенезі
заміняються іншими, обхідними. Таких
обхідних реакцій при глюкогенезі є три.
Перша – це синтез фосфоенолпірувату
із пірувату. Вона відбувається з
утворенням проміжного продукту –
оксалоацетату шляхом карбоксилювання
пірувату та його наступного фосфорилювання
і декарбоксилювання з утворенням
фосфоенолпірувату:
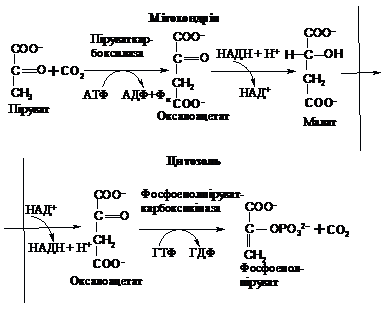 Як
видно із схеми, енергетичні витрати
цього процесу становлять дві макроергічні
фосфатні групи (одна – за рахунок ГТФ
і одна – за рахунок АТФ). Слід відзначити,
що дві стадії цієї реакції відбуваються
в різних компартментах: перша – в
мітохондріях, а друга – в цитозолі. В
мітохондріях оксалоацетат зазнає
відновлення за участю НАДН до малату
(яблучної кислоти), який транспортується
через мембрану в цитозоль, там знову
окиснюється до оксалоацетату за участю
НАД+.
Дві наступні обхідні реакції
пов’язані з дефосфорилюванням
фруктозо-1,6-дифосфату та глюкозо-6-фосфату.
Ці перетворення відбуваються шляхом
гідролізу з вивільненням фосфату.
Сумарний енергетичний ефект глюкогенезу
становить –6 макроергічних фосфатних
груп на одну молекулу глюкози, тобто
витрати на синтез є більшими ніж
енергетичний виграш при окисненні
молекули.
Сумарне рівняння глюкогенезу
можна записати так:
2С3Н4О3+4АТФ+2ГТФ+2НАДН+2Н+
® С6Н12О6+4АДФ+2ГДФ+6Фн+2НАД+
Весь хід
перетворень, які здійснюються при
глюкогенезі, представлений на
схемі:
Як
видно із схеми, енергетичні витрати
цього процесу становлять дві макроергічні
фосфатні групи (одна – за рахунок ГТФ
і одна – за рахунок АТФ). Слід відзначити,
що дві стадії цієї реакції відбуваються
в різних компартментах: перша – в
мітохондріях, а друга – в цитозолі. В
мітохондріях оксалоацетат зазнає
відновлення за участю НАДН до малату
(яблучної кислоти), який транспортується
через мембрану в цитозоль, там знову
окиснюється до оксалоацетату за участю
НАД+.
Дві наступні обхідні реакції
пов’язані з дефосфорилюванням
фруктозо-1,6-дифосфату та глюкозо-6-фосфату.
Ці перетворення відбуваються шляхом
гідролізу з вивільненням фосфату.
Сумарний енергетичний ефект глюкогенезу
становить –6 макроергічних фосфатних
груп на одну молекулу глюкози, тобто
витрати на синтез є більшими ніж
енергетичний виграш при окисненні
молекули.
Сумарне рівняння глюкогенезу
можна записати так:
2С3Н4О3+4АТФ+2ГТФ+2НАДН+2Н+
® С6Н12О6+4АДФ+2ГДФ+6Фн+2НАД+
Весь хід
перетворень, які здійснюються при
глюкогенезі, представлений на
схемі:
 Продукти
аеробного окиснення глюкози активують
процеси глюкогенезу і глікогенезу і
пригнічують дальший розпад глюкози.
Вже згадувалось, що в ролі таких
алостеричних регуляторів виступають
ацетил-КоА, лимонна кислота, АТФ. Таким
чином, напрямок перетворення вуглеводів
визнача
class=Section19>
ється
п
class=Section20>
отребами
організму.
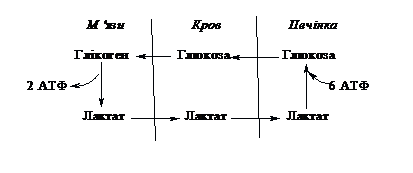
Існує взаємозв’язок між
депо вуглеводів (печінкою) і працюючими
органами (м’язами) в забезпеченні
повторного використання ресурсів. При
посиленому розпаді глікогену під час
роботи в м’язах в кров виділяється
молочна кислота. З током крові вона
попадає в клітини печінки. Там з неї
шляхом глюкогенезу синтезуються глюкоза
і глікоген. При потребі глікоген печінки
розпадається до глюкози, яка виходить
в кров, транспортується в м’язи і
забезпечує там ресинтез глікогену для
виконання нового навантаження. В клітинах
м’язів реакції глюкогенезу
class=Section21>
не
відбуваються. Описаний процес називається
циклом Корі:
Продукти
аеробного окиснення глюкози активують
процеси глюкогенезу і глікогенезу і
пригнічують дальший розпад глюкози.
Вже згадувалось, що в ролі таких
алостеричних регуляторів виступають
ацетил-КоА, лимонна кислота, АТФ. Таким
чином, напрямок перетворення вуглеводів
визнача
class=Section19>
ється
п
class=Section20>
отребами
організму.
Існує взаємозв’язок між
депо вуглеводів (печінкою) і працюючими
органами (м’язами) в забезпеченні
повторного використання ресурсів. При
посиленому розпаді глікогену під час
роботи в м’язах в кров виділяється
молочна кислота. З током крові вона
попадає в клітини печінки. Там з неї
шляхом глюкогенезу синтезуються глюкоза
і глікоген. При потребі глікоген печінки
розпадається до глюкози, яка виходить
в кров, транспортується в м’язи і
забезпечує там ресинтез глікогену для
виконання нового навантаження. В клітинах
м’язів реакції глюкогенезу
class=Section21>
не
відбуваються. Описаний процес називається
циклом Корі:
 Глюкогенез
є вторинним синтезом
глюкози, оскільки для нього використовуються
органічні молекули, вже синтезовані
живими організмами. На відміну від
нього, утворення глюкози в процесі
фотосинтезу з неорганічних речовин
(СО2 і Н2О) називається первинним синтезом.
Цей
процес можливий завдяки акцептуванню
енергії Сонця в АТФ і НАДФ:
АДФ + Фн +
hn ® АТФ + Н2О (фотофосфорилювання)
2Н2О
+ 4НАДФ+ ® 4НАДФН + 4Н+ + О2 (відновлення в
окисно-відновному ланцюгу хлоропластів)
АТФ
використовується при фотосинтезі для
активації рибулозо-5-фосфату. В цій
реакції утворюється рибулозо-1,5-дифосфат,
який і є акцептором СО2:
Глюкогенез
є вторинним синтезом
глюкози, оскільки для нього використовуються
органічні молекули, вже синтезовані
живими організмами. На відміну від
нього, утворення глюкози в процесі
фотосинтезу з неорганічних речовин
(СО2 і Н2О) називається первинним синтезом.
Цей
процес можливий завдяки акцептуванню
енергії Сонця в АТФ і НАДФ:
АДФ + Фн +
hn ® АТФ + Н2О (фотофосфорилювання)
2Н2О
+ 4НАДФ+ ® 4НАДФН + 4Н+ + О2 (відновлення в
окисно-відновному ланцюгу хлоропластів)
АТФ
використовується при фотосинтезі для
активації рибулозо-5-фосфату. В цій
реакції утворюється рибулозо-1,5-дифосфат,
який і є акцептором СО2:
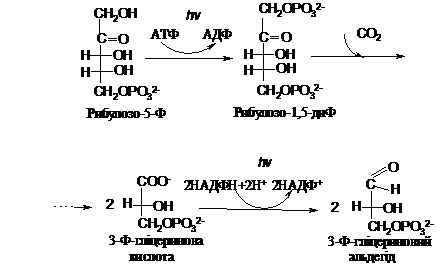 В
результаті приєднання СО2 до
рибулозо-1,5-дифосфату утворюється
нестійка проміжна сполука, яка містить
шість атомів С. Вона легко розпадається
на дві молекули 3-фосфогліцеринової
кислоти (3-ФГ). Для відновлення цієї
кислоти в 3-фосфогліцериновий альдегід
(3-ФГА) використовується НАДФН – продукт
фотовідновлення. Далі реакції синтезу
відбуваються аналогічно до
глюкогенезу.
Можна
зауважити, що реакції карбоксилювання,
в яких СО2 акцептується органічною
речовиною відбуваються як у первинному,
так і вторинному синтезі глюкози. Однак
продукт таких реакцій у вторинних
синтезах легко декар-боксилюється,
відщеплюючи знову молекулу СО2. На
відміну від вторинного, в первинному
синтезі відбувається реальне збільшення
вуглецевого ланцюга молекули за рахунок
карбону СО2 і реальне відновлення
утворених сполук за рахунок енергії
Сонця, а не окиснення інших органічних
субстратів.
Глюкозо-6-фосфат, продукт
фотосинтезу, є попередником інших
вуглеводів - сахарози, крохмалю, целюлози,
які синтезуються тільки в рослинних
клітинах. В перетворенні глюкозофосфату
в інші вуглеводи беруть участь
нуклеозидфосфати УТФ, ГТФ, АТФ.
Таким
чином, вуглеводи є первинними органічними
продуктами, які утворюються з неорганічних
за рахунок енергії світла фотосинтезуючими
організмами. Для інших організмів вони
є легко окиснюваними джерелами енергії,
які можуть забезпечувати роботу як в
анаеробних процесах (гліколіз), так і
при аеробному окисненні.
Багато
метаболітів, які утворюються в обміні
вуглеводів, можуть використовуватись
в обміні інших речовин.
В
результаті приєднання СО2 до
рибулозо-1,5-дифосфату утворюється
нестійка проміжна сполука, яка містить
шість атомів С. Вона легко розпадається
на дві молекули 3-фосфогліцеринової
кислоти (3-ФГ). Для відновлення цієї
кислоти в 3-фосфогліцериновий альдегід
(3-ФГА) використовується НАДФН – продукт
фотовідновлення. Далі реакції синтезу
відбуваються аналогічно до
глюкогенезу.
Можна
зауважити, що реакції карбоксилювання,
в яких СО2 акцептується органічною
речовиною відбуваються як у первинному,
так і вторинному синтезі глюкози. Однак
продукт таких реакцій у вторинних
синтезах легко декар-боксилюється,
відщеплюючи знову молекулу СО2. На
відміну від вторинного, в первинному
синтезі відбувається реальне збільшення
вуглецевого ланцюга молекули за рахунок
карбону СО2 і реальне відновлення
утворених сполук за рахунок енергії
Сонця, а не окиснення інших органічних
субстратів.
Глюкозо-6-фосфат, продукт
фотосинтезу, є попередником інших
вуглеводів - сахарози, крохмалю, целюлози,
які синтезуються тільки в рослинних
клітинах. В перетворенні глюкозофосфату
в інші вуглеводи беруть участь
нуклеозидфосфати УТФ, ГТФ, АТФ.
Таким
чином, вуглеводи є первинними органічними
продуктами, які утворюються з неорганічних
за рахунок енергії світла фотосинтезуючими
організмами. Для інших організмів вони
є легко окиснюваними джерелами енергії,
які можуть забезпечувати роботу як в
анаеробних процесах (гліколіз), так і
при аеробному окисненні.
Багато
метаболітів, які утворюються в обміні
вуглеводів, можуть використовуватись
в обміні інших речовин.