

MFK_2014_osen_Globalnie_izmeneniya_2
.pdfГлобальные изменения, кризисы и катастрофы: проблемы и достижения современной экологии
Лекция 2
Способна ли экология к прогнозу?
Сравнительно-видовые макроэкологические зависимости и связи между ними
Макроэкология
Макроэкология изучает экологические явления и процессы
(1)протекающие на больших пространствах,
(2)охватывающие длительные времена и
(3)включающие большие совокупности видов.
Большие пространства:
пространства, сравнимые по размерам с размерами континентов или океанических акваторий.
Большие времена:
времена, сравнимые с длительностью эволюции большой таксономической группы (например, млекопитающих).
Большие совокупности видов:
совокупность видов, населяющих большие пространства, или совокупность представителей большой таксономической группы. В последнем случае макроэкология выступает как
сравнительно-видовая экология
См. Л.В. Полищук. ТРОИЦКИЙ ВАРИАНТ, 2011, № 19 (88), с.12
Противопоставление популяционной экологии и макроэкологии не отрицает возможности создания теории экологических явлений как в локальном, так и, тем более, в глобальном масштабе.
Она лишь пытается расставить приоритеты:
поиск (более или менее) универсальных
инструментов
в локальном масштабе и
поиск (более или менее) универсальных зависимостей
в глобальном масштабе
Материалом для макроэкологии, как и других разделов экологии, являются данные по биологии отдельных видов

Стеллерова корова
(Hydrodamalis gigas)
Drawing thought to be the only remaining illustration of a dead female examined by Steller, 1743. Many later depictions were based on it.
Источник: https://en.wikipedia.org/wiki/ Steller%27s_sea_cow
Млекопитающее из семейства дюгоневых, эволюционно родственники современных слонов.
Обитала на Командорских островах в Тихом океане.
Крупное растительноядное животное («корова»!)
Была открыта в 1741 г., а уже в 1768 г., всего
через 27 лет, полностью истреблена.
Причины истребления:
Узкий ареал
Малая подвижность
Крупный размер
Низкая скорость размножения. Точные цифры неизвестны (не успели изучить), но наверняка она была низкой, как у всех крупных зверей.

Птица додо (маврикийский дронт) (Raphus cucullatus)
Источник: http://www.zoopicture.ru/dront/
Птица подсемейства дронтов, эволюционно родственники современных голубей.
Эндемик острова Маврикий в Индийском океане.
Последнее прижизненное наблюдение – 1662 год.
Причины истребления:
Узкий ареал (островной вид!)
Малая подвижность (не летали, а только бегали по земле)
Крупный размер (высотой около 1 м)
Низкая скорость размножения (в кладке было одно яйцо).
Среди макроэкологических зависимостей нас прежде всего интересуют аллометрические зависимости
Аллометрические зависимости
Аллометрические зависимости – это сравнительно-видовые зависимости экологических характеристик от массы тела. В качестве экологических характеристик в этом контексте рассматривают среднюю продолжительность жизни особи, скорость развития и размножения, численность (плотность) популяции и
даже риск вымирания. К базовым экологическим характеристикам
традиционно относят и среднюю скорость потребления кислорода (скорость основного обмена), хотя это скорее физиологическая, а не экологическая характеристика. «Основной» в выражении «основной обмен» означает скорость потребления кислорода в состоянии покоя, то есть когда физическая активность организма минимальна. При этом энергия, выделяющаяся при окислении органических веществ, поступающих с пищей, расходуется только на поддержание жизнедеятельности (дыхание, кровообращение, переваривание пищи, деятельность мозга)

Крупные организмы потребляют больше кислорода при дыхании (1)
Зависимость скорости основного обмена R (кДж/сут) от массы тела W (кг) для млекопитающих и птиц («кривая от мыши до слона»)
R = 288 W0.76
Источник : Гильманов Г.Г. 1987. Введение в количественную трофологию и экологическую биоэнергетику наземных позвоночных, Рис. 4
Исходный источник: Benedict F.G. 1938. Vital Energetics: A Study in Comparative Basal Metabolism

Техническое дополнение: Оксикалорийный коэффициент
Оксикалорийный коэффициент – это количество энергии, выделяющееся из того количества органического вещества пищи, на окисление
которого расходуется 1 г (или 1 мл) кислорода. Оксикалорийный коэффициент выражается в ккал/г O2 или кДж/г O2. На полное окисление 1 г белка тратится 1.748 г О2. Поскольку калорийность 1 г белка (то есть энергия, освобождающаяся при его окислении) составляет 5.65 ккал, оксикалорийный коэффициент для белка равен 5.65/1.748 = 3.23 ккал/г O2 или 13.6 кДж/г O2. Средний оксикалорийный коэффициент (с учетом содержания в организме белков, жиров и углеводов) принимают равным 3.38 ккал/г O2. Эта величина оксикалорийного коэффициента
была впервые рассчитана В.С. Ивлевым в 1934 г. (Ivlev 1934) и до сих пор широко
используется в экологии (Алимов 1989).
Соотношения между единицами измерения энергии: 1 Ккал = 4.2 кДж, 1 кДж = 239 кал
См. А.Ф. Алимов. Введение в продукционную гидробиологию. 1989, с. 13. А.Ф. Алимов приводит значение оксикалорийного коэффициента 14.2 кДж/г O2, что эквивалентно 3.38 ккал/г O2 (3.38 * 4.2 = 14.2).
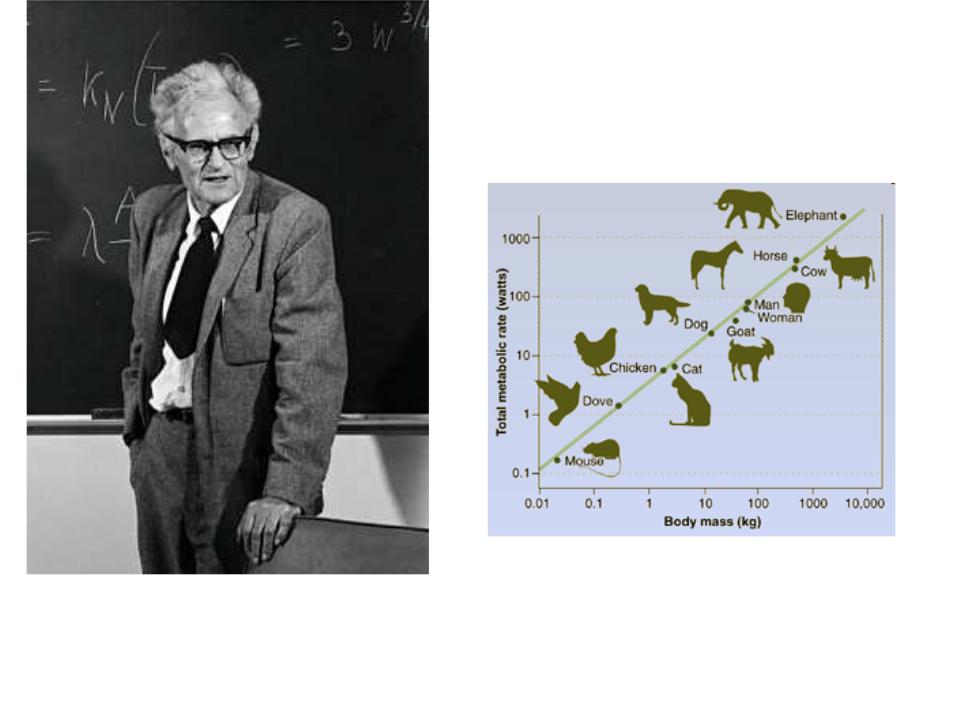
Фото: University of California, Davis http://books.nap.edu/openbook.php?record_id=11634&page=52 Из лекций А.М. Гилярова, с изменениями