

биохимия 3
.docБиохимия № 3Нуклеиновые кислоты
НК – ДНК и РНК
СОСТАВ: НК(полинуклеотиды)![]()
ДНК РНК
Н3РO4 Н3РO4
Дезоксирибоза Рибоза
Аденин Аденин
Гуанин Гуанин
Цитозин Цитозин
Тимин Урацил
Все АО можно разделить на пиримидиновые и пуриновые.
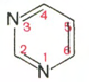 пиримидин
пиримидин
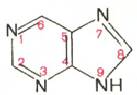 пурин
пурин
В составе НК встречаются пиримидиновых основания:
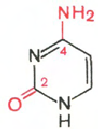 цитозин
цитозин
 урацил
урацил
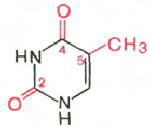 Тимин
Тимин
Пуриновых основания:
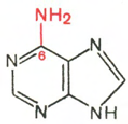 аденин
аденин
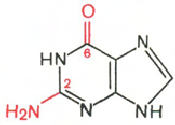 гуанин
гуанин
Углеводы в находятся в β-D-рибофуранозной форме:
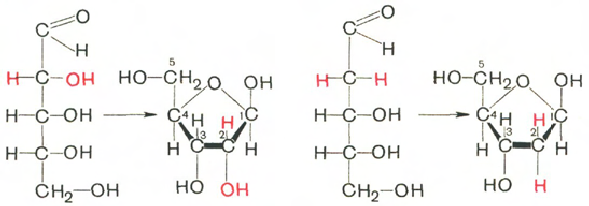
D-рибоза β-D-рибофураноза D-2-дезоксирибоза β-D-2-дезокси-рибофураноза
Нуклеозиды – АО+углевод (рибоза или дезоксирибоза)
Образование: у пиримидиновых АО отрывается Н от 1 атома N, у пуриновых – Н от 9 атома N и гликозидный гидроксил от углевода.
Мононуклеотиды – АО + углевод +Н3РО4
Н3РО4 присоединяется к нуклеозиду в положение 3’ или 5’ с образованием фосфоэфирной связи.
Подобно белкам, нуклеиновые кислоты — биополимеры, а их функция заключается в хранении, реализации и передаче генетической (наследственной) информации в живых организмах.
Существует два типа нуклеиновых кислот — дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Мономерами в нуклеиновых кислотах служат нуклеотиды. Каждый из них содержит азотистое основание, пятиуглеродный сахар (дезоксирибоза — в ДНК, рибоза — в РНК) и остаток фосфорной кислоты.
В ДНК входят четыре вида нуклеотидов, отличающихся по азотистому основанию в их составе, — аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т). В молекуле РНК также имеется 4 вида нуклеотидов с одним из азотистых оснований — аденином, гуанином, цитозином и урацилом (У). Таким образом, ДНК и РНК различаются как по содержанию сахара в нуклеотидах, так и по одному из азотистых оснований.
Молекулы ДНК и РНК существенно различаются по своему строению и выполняемым функциям.
Молекула ДНК может включать огромное количество нуклеотидов — от нескольких тысяч до сотен миллионов. В структурном отношении она представляет собой двойную спираль из полинуклеотидных цепей, соединенных с помощью водородных связей между азотистыми основаниями нуклеотидов. Благодаря этому полинуклеотидные цепи прочно удерживаются одна возле другой.
При исследовании различных ДНК было установлено, что аденин одной цепи может связываться лишь с тимином, а гуанин — только с цитозином другой. Следовательно, порядок расположения нуклеотидов в одной цепи строго соответствует порядку их расположения в другой. Этот феномен получил название комплементарности (т. е. дополнения), а противоположные полинуклеотидные цепи называются комплементарными. Именно этим обусловлено уникальное среди всех неорганических и органических веществ свойство ДНК — способность к самовоспроизведению или удвоению. При этом сначала комплементарные цепи молекул ДНК расходятся (под воздействием специального фермента происходит разрушение связей между комплементарными нуклеотидами двух цепей). Затем на каждой цепи начинается синтез новой («недостающей») комплементарной ей цепи за счет свободных нуклеотидов, всегда имеющихся в большом количестве в клетке. В результате вместо одной («материнской») молекулы ДНК образуются две («дочерние») новые, идентичные по структуре и составу друг другу, а также исходной молекуле ДНК. Этот процесс всегда предшествует клеточному делению и обеспечивает передачу наследственной информации от материнской клетки дочерним и всем последующим поколениям.
Различают 3 уровня структурной организации ДНК:
Первичная структура – это спирально изогнутая полинуклеотидная цепь с определенным качественным и количественным набором мононуклеотидов, которые связаны 3’5’-фосфодиэфирной связью Т.о., каждая цепь имеет 5’конец (фосфатный) и 3’конец (дезоксирибоза). Участки ДНК, содержащие генетическую информацию, называются структурными генами.
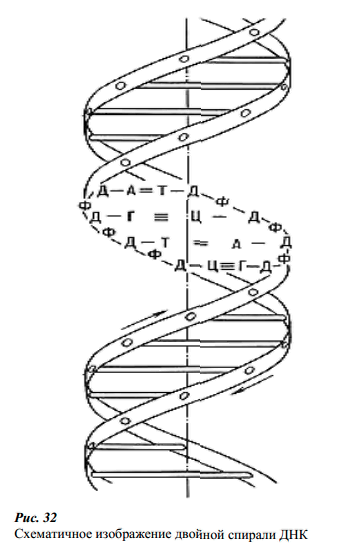 Вторичная структура – это
двухспиральная молекула, полинуклеотидные
цепи которой антипараллельны и связаны
водородными связями между комплементарными
основаниями обоих цепей. Один виток
спирали содержит 10 нуклеотидных остатков.
Вторичная структура – это
двухспиральная молекула, полинуклеотидные
цепи которой антипараллельны и связаны
водородными связями между комплементарными
основаниями обоих цепей. Один виток
спирали содержит 10 нуклеотидных остатков.
1. Молекулы ДНК построены из двух полинуклеотидных цепей, ориентированных антипараллельно и по всей длине связанных друг с другом водородными связями (причем в образовании водородных связей участвует каждый мононуклеотид).
2. Водородные связи между цепями образуются за счет специфических взаимодействий остатка аденина одной цепи с остатком тимина другой цепи (пара А…Т) и остатка гуанина одной цепи с остатком цитозина другой цепи (пара Г…Ц). Образование водородных связей в парах А…Т и Г…Ц.
3. Первичная структура одной цепи молекулы ДНК в составе двойной цепи комплементарна первичной структуре другой цепи. Это положение легко понять на примере следующей схемы:
Если в положении п (считая с 5'-конца) первой цепи находится остаток аденина, то в положении п (считая с З'-конца) второй цепи находится комплементарный ему остаток тимина, а не другое азотистое основание.
4. Обе цепи закручены в спираль — двойная спираль или дуплекс, которая имеет общую ось . При этом цепи могут быть разделены только путем раскручивания (такие спирали называют плектонемическими). Азотистые основания обращены внутрь спирали; их плоскости перпендикулярны оси спирали и параллельны друг другу, образуя внутри спирали стопку оснований. Между основаниями в этой стопке возникают гидрофобные, или стэкинг-взаимодействия, которые, наряду с водородными связями, способствуют стабилизации структуры двойной спирали в пространстве. Азотистые основания упакованы очень плотно и не контактируют с окружающим водным раствором.
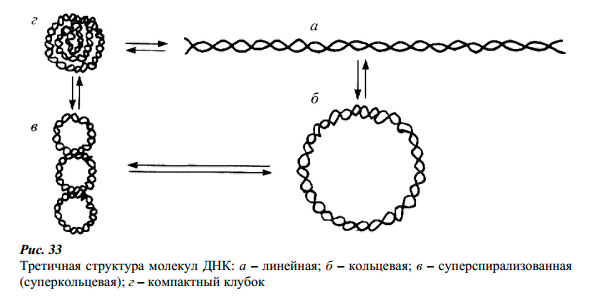 Третичная
структура. В частицах вирусов, клетках
бактерий и высших организмов, молекулы
ДНК плотно «упакованы» и образуют
довольно сложные структуры. Вирусную
ДНК можно отнести к сравнительно мелким
полимерным биомолекулам, но если ее
вытянуть, то она окажется во много раз
длиннее, чем сам вирус. Выделенные из
вирусных частиц молекулы ДНК имеют либо
линейную, либо кольцевую форму.
Линейные молекулы ДНК in
vivo свертываются
в плотный клубок. В таком состоянии
они более устойчивы к деградации.
Кольцевую ковалентно-связанную структуру
имеют двухцепочечные ДНК бактерий,
вирусов, плазмид, митохондрий и др.
Двухцепочечные кольцевые ДНК легко
переходят в суперспирализованное
состояние, которое обеспечивает более
плотную упаковку громадной молекулы
ДНК в малом объеме ядра или клетки.
Третичная
структура. В частицах вирусов, клетках
бактерий и высших организмов, молекулы
ДНК плотно «упакованы» и образуют
довольно сложные структуры. Вирусную
ДНК можно отнести к сравнительно мелким
полимерным биомолекулам, но если ее
вытянуть, то она окажется во много раз
длиннее, чем сам вирус. Выделенные из
вирусных частиц молекулы ДНК имеют либо
линейную, либо кольцевую форму.
Линейные молекулы ДНК in
vivo свертываются
в плотный клубок. В таком состоянии
они более устойчивы к деградации.
Кольцевую ковалентно-связанную структуру
имеют двухцепочечные ДНК бактерий,
вирусов, плазмид, митохондрий и др.
Двухцепочечные кольцевые ДНК легко
переходят в суперспирализованное
состояние, которое обеспечивает более
плотную упаковку громадной молекулы
ДНК в малом объеме ядра или клетки.
Роль ДНК: 1) хранение и передача наследственной информации; 2) биосинтез ферментов, белков и гормонов.
Молекулы РНК, как правило, одноцепочечные (в отличие от ДНК) и содержат значительно меньшее число нуклеотидов. Выделяют три вида РНК, различающиеся по величине молекул и выполняемым функциям, — информационную или матричную (и-РНК), рибосомальную (р-РНК) и транспортную (т-РНК).
Информационная РНК (и-РНК) Эта РНК не имеет жесткой специфической структуры и ее полинуклеотидная цепь образует изогнутые петли. В нерабочем состоянии и-РНК собрана в складки, свернута в клубок, связана с белком; а во время функционирования цепь расправляется. Матричные РНК синтезируются на ДНК в ядре. Процесс называется транскрипция (списывание). Роль м-РНК – она несет информацию об аминокислотной последовательности (т.е. первичной структуры) синтезируемого белка. Место каждой аминокислоты в молекуле белка закодировано определенной последовательностью нуклеотидов в цепи м-РНК, т.е. в м-РНК имеются «кодовые слова» для каждой аминокислоты – триплеты, или кодоны, или генетические коды. Свойства генетического кода: 1) триплетность. Из 4-х возможных мононуклеотидов м-РНК (УМФ, ГМФ, АМФ, ЦМФ) можно построить по правилам перестановки 64 кодона. 61 кодон шифрует 20 аминокислот, а 3 кодона (УАА, УАГ, УГА) не кодируют ни одной аминокислоты. Они играют роль терминирующих (или «стоп-кодонов»), т.к. на них останавливается синтез п/п цепи; 2) неперекрещиваемость – списывание информации идет только в одном направлении; 3) непрерывность – код является линейным, однонаправленным; 4) универсальность, т.е. одна и та же аминокислота у всех живых организмов кодируется одинаковыми кодами у всех живых существ; 5) вырожденность. Первые две буквы кодона определяют его специфичность, третья менее специфична. Известно 20 аминокислот, а кодонов 61, следовательно, большинство аминокислот кодируется несколькими кодонами (2-6).
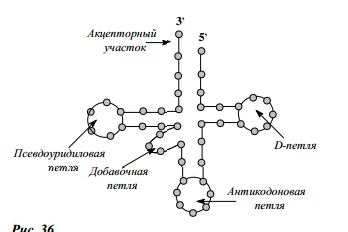
Транспортная РНК (т-РНК).Содержится в цитоплазме, мол.масса небольшая состоит из 70-80 нуклеотидов. Основная роль – транспорт и установка аминокислот на комплиментарном кодоне м-РНК. т-РНК специфичны к аминокислотам, что обеспечивается ферментом аминоацилсинтетазой. В неактивном состоянии она свернута в клубочек, а в активном имеет вид трилистника (клеверного листа). В молекуле т-РНК различают несколько участков: а) акцепторный стебель с последовательностью нуклеотидов АЦЦ, к нему присоединяется аминокислота. Б) участок для присоединения к рибосоме; в) антикодон – участок, комплиментарный кодону м-РНК, который кодирует аминокислоту, присоединенную к данной т-РНК. Особенностью первичной структуры т-РНК является то, что они содержат минорные, или модифицированные основания, которые способны к неклассическому спариванию. Это ускоряет белковый синтез. Т.о., т-РНК «метит» аминокислоту, придавая ей специфичность и способствует установлению аминокислоты на определенный участок м-РНК.
 Рибосомальная
РНК (р-РНК) содержится в ядрышке и
рибосомах клетки, имеет цепь средней
длины. Одна р-РНК способна соединяться
с 30 молекулами белка. Полинуклеотидная
цепь р-РНК легко изгибается и укладывается
вместе с белком в компактные тельца.
Рибосома состоит из 2-х субъдиниц –
большой и малой. В рибосоме различают
2 участка – А (аминокислотный, или участок
узнавания) и Р – пептидный, здесь
присоединяется п/п цепь. Эти центры
расположены на контактирующих поверхностях
обеих субъдиниц. Рибосомы могут свободно
перемещаться в клетке, что дает возможность
синтезировать белки в клетке там, где
это необходимо. Рибосомы мало специфичны
и могут считывать информацию с чужеродных
м-РНК, вместе с м-РНК рибосомы образуют
матрицу. Роль р-РНК – обуславливает
количество синтезируемого белка.
Рибосомальная
РНК (р-РНК) содержится в ядрышке и
рибосомах клетки, имеет цепь средней
длины. Одна р-РНК способна соединяться
с 30 молекулами белка. Полинуклеотидная
цепь р-РНК легко изгибается и укладывается
вместе с белком в компактные тельца.
Рибосома состоит из 2-х субъдиниц –
большой и малой. В рибосоме различают
2 участка – А (аминокислотный, или участок
узнавания) и Р – пептидный, здесь
присоединяется п/п цепь. Эти центры
расположены на контактирующих поверхностях
обеих субъдиниц. Рибосомы могут свободно
перемещаться в клетке, что дает возможность
синтезировать белки в клетке там, где
это необходимо. Рибосомы мало специфичны
и могут считывать информацию с чужеродных
м-РНК, вместе с м-РНК рибосомы образуют
матрицу. Роль р-РНК – обуславливает
количество синтезируемого белка.
Все виды РНК образуются в процессе транскрипции соответствующих генов ДНК.