

Характеристика репликации.
Субстратами для репликации служат дезоксирибонуклеозидфосфаты (дАТФ, дЦТФ, дГТФ, дТТФ).
Источники энергии - дАТФ, дГТФ, дТТФ, дЦТФ.
Репликация ДНК всегда идет с образованием репликативной вилки, синтез идет в направлении 5—»3. В синтезе принимает участие мультиферментный репликативный комплекс.
Ферменты: эндонуклеазы, ДНК - раскручивающие белки, ДНК-полимеразы (а,δ,β, ε), ДНК - лигаза, РНК - праймаза.
Продукты реакции - вновь синтезированные молекулы ДНК идентичны матрице, репликация идет в двух направлениях, обе цепи реплицируются одновременно, протекает в S фазу клеточного цикла. Полная репликация генома происходит за 9 часов -время необходимое для удвоения генетического материала диплоидной делящейся клетки —> репликация начинается в нескольких местах с образованием «репликативного глаза».
Если точка репликации находится в начале «репликативного глаза», то репликация идет в одном направлении.
Если точка репликации находится в середине «репликативного глаза», то репликация идет в двух направлениях, но цифровое сохраняется.
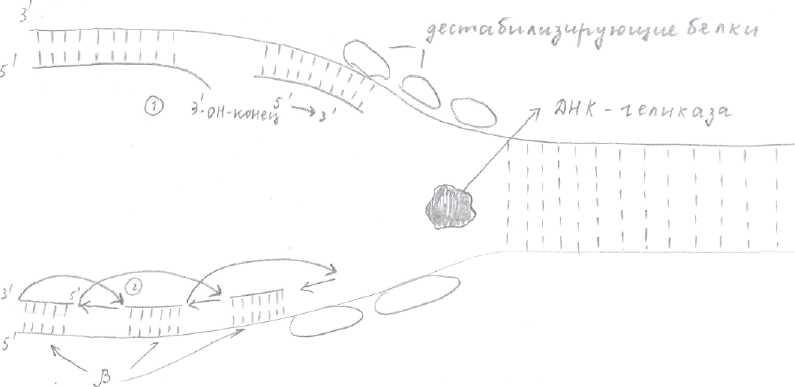
Образование репликативных вилок начинается с образования «репликативного глаза». Для бактерий и вирусов «репликативный глаз» образуется там, где имеются специфические нуклеотидные последовательности (около 300 нуклеотидов) - это точки начала репликации. В точке начала репликации эндонуклеаза (топоизомераза) вызывает кратковременный разрыв 1 цепи ДНК (это топоизомераза 1) или двухцепочечный разрыв (топоизомераза 2) и присоединяется к разорванному концу, способствуя расплетению небольшого участка спирали. В дальнейшем раскручивании ДНК принимают участие спецефические белки - ДНК-геликазы (рис.2), использующие АТФ (АТФ-зависимые ДНК-геликазы).
Другие белки образуют репликативную вилку - дестабилизирующие белки (рис.2), которые выстраиваются в ряды на образовавшейся одноцепочечной ДНК, растягивают её остов и делают основания нуклеотидов более доступными для спаривания. Для того чтобы репликативная вилка могла продвигаться вперёд со скоростью равной 50 нуклеотидов в секунду, родительская спираль должна вращаться 50 оборотов в секунду. Этого не происходит, т.к. при действии топоизомеразы образуется шарнирный участок, т.е. временный разрыв цепи, возникающий под её действием, даёт возможность спирали ДНК по обе стороны от него вращаться.
Синтез, растущей дочерней цепи идёт в направлении 5—>3, осуществляется ДНК-полимеразой (ДНК-полимераза 5 осуществляет синтез лидирующей цепи, е, а, р -отстающей), осуществляющей полимеризацию нуклеотидов по принципу комплиментарности. Рис.2
Ведущая цепь синтезируется следующим образом:
Синтез начинается с синтеза затравки (т.к. ДНК-полимераза может только достраивать нуклеотиды к имеющимся 3-ОН-концам). Затравка или праймер (от 10 до 200 нуклеотидов) состоит из нуклеотидов РНК и синтезируется РНК-праймазой. 3-ОН-конец праймера должен быть спарен с матричной цепью. Если 3-ОН-конец не спарен с нуклеотидами матричной цепи, тогда ДНК-полимеразы отщепляет неспаренные нуклеотиды до появления спаренного конца (1) (рис.2).
ДНК-полимераза 5 достраивает дочернюю цепь, прикрепляя к праймеру 5-конец (2). Синтез на ведущей цепи протекает быстро. Синтез отстающей цепи происходит особенным образом, т.к. если бы он проходил также, как на ведущей цепи, направление синтеза было бы 3—>5 (рис.2), а ДНК-полимеразы осуществляет "синтез новой дочерней цепи в направлении 5^------>3^ —> особенности: - отстающая дочерняя цепь синтезируется фрагментами (5;
для каждого фрагмента нужен свой праймер. От каждого праймера синтезируются в нужном направлении фрагменты от 1000 до 2000 нуклеотидов у прокариот, у эукариот - 100-200- фрагменты Оказаки.
для синтеза фрагментов Оказаки ДНК-полимеразе (а) необходимо ~ 4с, после этого она осуществляет синтез следующего фрагмента.
Каждый раз нужна новая затравка (из 10 нуклеотидов). Затравки синтезируются с определенным интервалом (3) на матрице, затем их наращивает ДНК-полимераза с 3- конца, начиная всякий раз новые фрагменты Оказаки, до тех пор, пока не достигнет РНК-затравки, присоединенной к 5-концу предыдущего фрагмента (4).
Затем ДНК-полимераза р удаляет затравки между фрагментами Оказаки и достраивает
уклеотидную цепь из нуклеотидов ДНК. Завершает процесс ДНК-лигаза, которая
соединяет З'-фрагмент нового фрагмента ДНК с 5-концом предыдущего.
Таким образом, в целом цепь строится в направлении 3^-»5,' но каждый фрагмент синтезируется в нужном направлении 5—»3 по типу шитья назад.