Гильдебрандт Александра
.docx
Кле́точная мембра́на (или
цитолемма, или плазмалемма, или
плазматическая мембрана) отделяет
содержимое любой клетки от внешней
среды, обеспечивая её целостность;
регулируют обмен между клеткой и средой;
внутриклеточные мембраны разделяют
клетку на специализированные замкнутые
отсеки — компартменты или органеллы,
в которых поддерживаются определённые
условия среды.
Мембраны
биологические (лат. membrana оболочка,
перепонка) функционально активные
поверхностные структуры толщиной в
несколько молекулярных слоев,
ограничивающие цитоплазму и большинство
органелл клетки, а также образующие
единую внутриклеточную систему канальцев,
складок, замкнутых областей.
Биологические мембраны имеются во всех
клетках. Их значение определяется
важностью функций, которые они выполняют
в процессе нормальной жизнедеятельности,
а также многообразием заболеваний и
патологических состояний, возникающих
при различных нарушениях мембранных
функций и проявляющихся практически
на всех уровнях организации — от клетки
и субклеточных систем до тканей, органов
и организма в целом.
Клеточная
стенка, если таковая у клетки имеется
(обычно есть у растительных клеток),
покрывает клеточную мембрану.
Клеточная мембрана представляет собой
двойной слой (бислой) молекул класса
липидов, большинство из которых
представляет собой так называемые
сложные липиды — фосфолипиды. Молекулы
липидов имеют гидрофильную («головка»)
и гидрофобную («хвост») часть. При
образовании мембран гидрофобные участки
молекул оказываются обращены внутрь,
а гидрофильные — наружу. Мембраны —
структуры инвариабельные, весьма сходные
у разных организмов. Некоторое исключение
составляют, пожалуй, археи, у которых
мембраны образованы глицерином и
терпеноидными спиртами. Толщина мембраны
составляет 7—8 нм.
Биологическая
мембрана включает и различные белки:
интегральные (пронизывающие мембрану
насквозь), полуинтегральные (погруженные
одним концом во внешний или внутренний
липидный слой), поверхностные (расположенные
на внешней или прилегающие к внутренней
сторонам мембраны). Некоторые белки
являются точками контакта клеточной
мембраны с цитоскелетом внутри клетки,
и клеточной стенкой (если она есть)
снаружи. Некоторые из интегральных
белков выполняют функцию ионных каналов,
различных транспортеров и
рецепторов.
Структура
и состав биомембран
Мембраны
состоят из липидов трёх классов:
фосфолипиды, гликолипиды и холестерол.
Фосфолипиды и гликолипиды
(липиды с присоединёнными к ним углеводами)
состоят из двух длинных гидрофобных
углеводородных «хвостов», которые
связаны с заряженной гидрофильной
«головой». Холестерол придаёт мембране
жёсткость, занимая свободное пространство
между гидрофобными хвостами липидов и
не позволяя им изгибаться. Поэтому
мембраны с малым содержанием холестерола
более гибкие, а с большим — более жёсткие
и хрупкие. Также холестерол служит
«стопором», препятствующим перемещению
полярных молекул из клетки и в клетку.
Важную часть мембраны составляют белки,
пронизывающие её и отвечающие за
разнообразные свойства мембран. Их
состав и ориентация в разных мембранах
различаются.
Клеточные мембраны
часто асимметричны, то есть слои
отличаются по составу липидов, переход
отдельной молекулы из одного слоя в
другой (так называемый флип-флоп)
затруднён.
Липиды
В составе
М.б. обнаружены липиды трех классов:
фосфолипиды, гликолипиды и стероиды. В
мембранах животных клеток более 50% всех
липидов составляют фосфолипиды —
глицерофосфолипиды (фосфатидилхолин,
фосфатидилэтаноламин, фосфатидилсерин,
фосфатидилинозит) и сфингофосфолипиды
(производные церамида, сфингомиелин).
Гликолипиды представлены цереброзидами,
сульфатидами и ганглиозидами, а стероиды
— в основном холестерином (около 30%). В
липидных компонентах М.б. содержатся
разнообразные жирные кислоты, однако
в мембранах животных клеток преобладают
пальмитиновая, олеиновая и стеариновая
кислоты. Основную структурную роль в
биологических мембранах играют
фосфолипиды. Они обладают выраженной
способностью формировать двухслойные
структуры (бислои) при смешивании с
водой, что обусловлено химической
структурой фосфолипидов, молекулы
которых состоят из гидрофильной части
— «головки» (остаток фосфорной кислоты
и присоединенная к нему полярная группа,
например холин) и гидрофобной части —
«хвоста» (как правило, две жирно-кислотные
цепи). В водной среде фосфолипиды бислоя
расположены таким образом, что
жирно-кислотные остатки обращены внутрь
бислоя и, следовательно, изолированы
от окружающей среды, а гидрофильные
«головки» —наоборот, наружу. Липидный
бислои представляет собой динамичную
структуру: образующие его липиды могут
вращаться, двигаться в латеральном
направлении и даже переходить из слоя
в слой (флип-флоп переход). Такое строение
липидного бислоя легло в основу
современных представлений о структуре
М.б. и определяет некоторые важные
свойства М.б., например способность
служить барьером и не пропускать молекулы
веществ, растворенных в воде (рис.).
Нарушение структуры бислоя может
привести к нарушению барьерной функции
мембран.
Холестерин в составе М.б.
играет роль модификатора бислоя, придавая
ему определенную жесткость за счет
увеличения плотности «упаковки» молекул
фосфолипидов.
Гликолипиды несут
разнообразные функции: отвечают за
рецепцию некоторых биологически активных
веществ, участвуют в дифференцировке
ткани, определяют видовую специфичность.
Белки биологических мембран исключительно
разнообразны. Молекулярная масса их в
большинстве своем составляет 25 000 — 230
000.
Белки могут взаимодействовать
с липидным бислоем за счет электростатических
и (или) межмолекулярных сил. Они
сравнительно легко могут быть удалены
из мембраны. К такому типу белков относят
цитохром с (молекулярная масса около
13 000), обнаруживаемый на наружной
поверхности внутренней мембраны
митохондрий.
Эти белки называются
периферическими, или наружными. Для
других белков, получивших название
интегральных, или внутренних, характерно
то, что одна или несколько полипептидных
цепей оказываются погруженными в бислои
или пересекают его, иногда не один раз
(например, гликофорин, транспортные
АТФ-азы, бактериородопсин). Часть белка,
контактирующая с гидрофобной частью
липидного бислоя, имеет спиральное
строение и состоит из неполярных
аминокислот, в силу чего между этими
компонентами белков и липидов происходит
гидрофобное взаимодействие. Полярные
группы гидрофильных аминокислот
непосредственно взаимодействуют с
примембранными слоями, как с одной, так
и с другой стороны бислоя. Молекулы
белков, как и молекулы липидов, находятся
в динамическом состоянии, для них также
характерна вращательная, латеральная
и вертикальная подвижность. Она является
отражением не только их собственной
структуры, но и функциональной активности.
что в значительной степени определяется
вязкостью липидного бислоя, которая, в
свою очередь, зависит от состава липидов,
относительного содержания и вида
ненасыщенных жирно-кислотных цепей.
Этим объясняется узкий температурный
диапазон функциональной активности
мембраносвязанных белков.
Белки
Белки мембран выполняют три основные
функции: каталитическую (ферменты),
рецепторную и структурную. Однако такое
разграничение достаточно условно, и в
ряде случаев один и тот же белок может
выполнять и репепторную и ферментную
функции (например, инсулин).
Число
мембранных ферментов (Ферменты) в клетке
достаточно велико, однако их распределение
в различных типах М.б. неодинаково.
Некоторые ферменты (маркерные) присутствуют
только в мембранах определенного типа
(например, Na, К-АТФ-аза, 5-нуклеотидаза,
аденилатциклаза — в плазматической
мембране; цитохром Р-450, НАДФН-дегидрогеназа,
цитохром в5 — в мембранах эндоплазматического
ретикулума; моноаминоксидаза — в
наружной мембране митохондрий, а цитохром
С-оксидаза, сукцинат-дегидрогеназа —
во внутренней; кислая фосфатаза — в
мембране лизосом).
Рецепторные
белки, специфически связывая
низкомолекулярные вещества (многие
гормоны, медиаторы), обратимо меняют
свою форму. Эти изменения запускают
внутри клетки ответные химические
реакции. Таким способом клетка принимает
различные сигналы, поступающие из
внешней среды.
К структурным белкам
относят белки цитоскелета, прилегающие
к цитоплазматической стороне клеточной
мембраны. В комплексе с микротрубочками
и микрофиламентами цитоскелета они
обеспечивают противодействие клетки
изменению ее объема и создают эластичность.
В эту же группу включают ряд мембранных
белков, функции которых не
установлены.
Углеводы
Углеводы в биологических мембранах
находятся в соединении с белками
(гликопротеины) и липидами (гликолипиды).
Углеводные цепи белков представляют
собой олиго- или полисахаридные структуры,
в состав которых входят глюкоза,
галактоза, нейраминовая кислота, фукоза
и манноза. Углеводные компоненты М.б.
открываются в основном во внеклеточную
среду, образуя на поверхности клеточных
мембран множество ветвистых образований,
являющихся фрагментами гликолипидов
или гликопротеидов. Их функции связаны
с контролем за межклеточным взаимодействием,
поддержанием иммунного статуса клетки,
обеспечением стабильности белковых
молекул в М.б. Многие рецепторные белки
содержат углеводные компоненты. Примером
могут служить антигенные детерминанты
групп крови, представленные гликолипидами
и гликопротеинами.
Мембранные
органеллы
Это замкнутые
одиночные или связанные друг с другом
участки цитоплазмы, отделённые от
гиалоплазмы мембранами. К одномембранным
органеллам относятся эндоплазматическая
сеть, аппарат Гольджи, лизосомы, вакуоли,
пероксисомы; к двумембранным — ядро,
митохондрии, пластиды. Строение мембран
различных органелл отличается по составу
липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают
избирательной проницаемостью: через
них медленно диффундируют глюкоза,
аминокислоты, жирные кислоты, глицерол
и ионы, причем сами мембраны в известной
мере активно регулируют этот процесс
— одни вещества пропускают, а другие
нет. Существует четыре основных механизма
для поступления веществ в клетку или
вывода их из клетки наружу: диффузия,
осмос, активный транспорт и экзо- или
эндоцитоз. Два первых процесса носят
пассивный характер, то есть не требуют
затрат энергии; два последних — активные
процессы, связанные с потреблением
энергии.
Избирательная
проницаемость мембраны при пассивном
транспорте обусловлена специальными
каналами — интегральными белками. Они
пронизывают мембрану насквозь, образовывая
своего рода проход. Для элементов K, Na и
Cl есть свои каналы. Относительно градиента
концентрации молекулы этих элементов
движутся в клетку и из неё. При раздражении
каналы натриевых ионов раскрываются,
и происходит резкое поступление в клетку
ионов натрия.
При этом происходит
дисбаланс мембранного потенциала. После
чего мембранный потенциал восстанавливается.
Каналы калия всегда открыты, через них
в клетку медленно попадают ионы калия.
Функции
Барьерная
— обеспечивает регулируемый, избирательный,
пассивный и активный обмен веществ с
окружающей средой. Например, мембрана
пероксисом защищает цитоплазму от
опасных для клетки пероксидов.
Избирательная проницаемость означает,
что проницаемость мембраны для различных
атомов или молекул зависит от их размеров,
электрического заряда и химических
свойств. Избирательная проницаемость
обеспечивает отделение клетки и клеточных
компартментов от окружающей среды и
снабжение их необходимыми веществами.
Транспортная — через мембрану
происходит транспорт веществ в клетку
и из клетки. Транспорт через мембраны
обеспечивает: доставку питательных
веществ, удаление конечных продуктов
обмена, секрецию различных веществ,
создание ионных градиентов, поддержание
в клетке оптимального pH и концентрации
ионов, которые нужны для работы клеточных
ферментов.
Частицы, по какой-либо
причине неспособные пересечь фосфолипидный
бислой (например, из-за гидрофильных
свойств, так как мембрана внутри
гидрофобна и не пропускает гидрофильные
вещества, или из-за крупных размеров),
но необходимые для клетки, могут
проникнуть сквозь мембрану через
специальные белки-переносчики
(транспортеры) и белки-каналы или путем
эндоцитоза.
При пассивном транспорте
вещества пересекают липидный бислой
без затрат энергии по градиенту
концентрации путем диффузии. Вариантом
этого механизма является облегчённая
диффузия, при которой веществу помогает
пройти через мембрану какая-либо
специфическая молекула. У этой молекулы
может быть канал, пропускающий вещества
только одного типа.
Активный транспорт
требует затрат энергии, так как происходит
против градиента концентрации. На
мембране существуют специальные
белки-насосы, в том числе АТФаза, которая
активно вкачивает в клетку ионы калия
(K+) и выкачивают из неё ионы натрия
(Na+).
Матричная — обеспечивает
определенное взаиморасположение и
ориентацию мембранных белков, их
оптимальное взаимодействие.
Механическая — обеспечивает
автономность клетки, ее внутриклеточных
структур, также соединение с другими
клетками (в тканях). Большую роль в
обеспечение механической функции имеют
клеточные стенки, а у животных —
межклеточное вещество.
Энергетическая
— при фотосинтезе в хлоропластах и
клеточном дыхании в митохондриях в их
мембранах действуют системы переноса
энергии, в которых также участвуют
белки;
Рецепторная — некоторые
белки, находящиеся в мембране, являются
рецепторами (молекулами, при помощи
которых клетка воспринимает те или иные
сигналы).
Например, гормоны,
циркулирующие в крови, действуют только
на такие клетки-мишени, у которых есть
соответствующие этим гормонам рецепторы.
Нейромедиаторы (химические вещества,
обеспечивающие проведение нервных
импульсов) тоже связываются с особыми
рецепторными белками клеток-мишеней.
Ферментативная — мембранные белки
нередко являются ферментами. Например,
плазматические мембраны эпителиальных
клеток кишечника содержат пищеварительные
ферменты.
Осуществление генерации
и проведения биопотенциалов. С помощью
мембраны в клетке поддерживается
постоянная концентрация ионов:
концентрация иона К+ внутри клетки
значительно выше, чем снаружи, а
концентрация Na+ значительно ниже, что
очень важно, так как это обеспечивает
поддержание разности потенциалов на
мембране и генерацию нервного импульса.
Маркировка клетки — на мембране
есть антигены, действующие как маркеры
— «ярлыки», позволяющие опознать клетку.
Это гликопротеины (то есть белки с
присоединенными к ним разветвленными
олигосахаридными боковыми цепями),
играющие роль «антенн». Из-за бесчисленного
множества конфигурации боковых цепей
возможно сделать для каждого типа клеток
свой особый маркер. С помощью маркеров
клетки могут распознавать другие клетки
и действовать согласованно с ними,
например, при формировании органов и
тканей. Это же позволяет иммунной системе
распознавать чужеродные антигены.
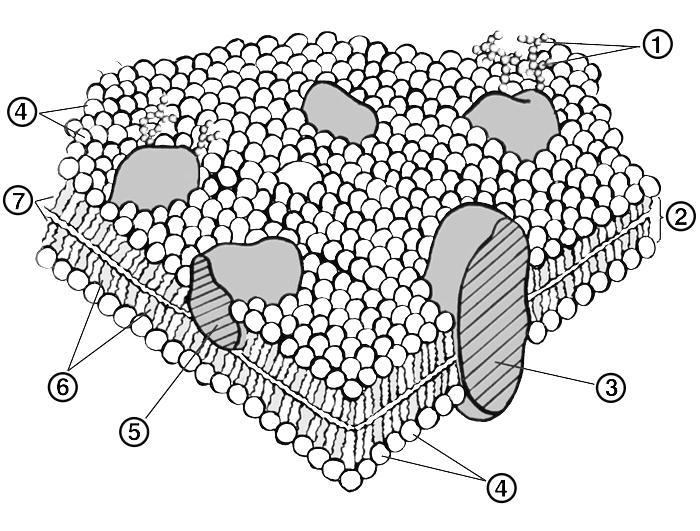
Схема
строения биологической мембраны:
1 — углеводные фрагменты гликопротеидов;
2 — липидный бислой; 3 — интегральный
белок; 4 — «головки» фосфолипидов; 5 —
периферический белок; 6 — холестерин;
7 — жирнокислотные «хвосты» фосфолипидов.