

2 курс / Нормальная физиология / Физиология ВНД
.pdfподготовил Дадаев Гаджимурад
ФИЗИОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Занятие №1.
1. Структурно-функциональная характеристика коры большого мозга
Кора является высшим отделом ЦНС, обеспечивающим на основе врожденных и приобретенных в онтогенезе функций наиболее совершенную организацию поведения организма.
Кора головного мозга представляет собой многослойную нейронную ткань с множеством складок общей площадью в обоих полушариях примерно 2200 см2, что соответствует квадрату со сторонами 47 х 47 см, ее объем соответствует 40 % массы головного мозга, ее толщина колеблется от 1,3 до 4,5 мм, а общий объем равен 600 см3. В состав коры головного мозга входит 109–1010 нейронов и множество глиальных клеток, общее число которых пока неизвестно.
Морфофункциональные особенности коры:
1)Многослойность расположения нейронов.
2)Модульный принцип организации.
3)Соматотопическая локализация рецепторных систем (т.е. каждый анализатор имеет свою проекцию в коре больших полушарий).
4)Экранный принцип проекции заключается в том, что рецептор проецирует свой сигнал не на один нейрон коры, а на их поле, которое образуется коллатералями и связями
нейронов. |
|
|
|
|
|
|
5) Зависимость |
уровня |
активности |
от |
влияния |
подкорковых |
структур |
и ретикулярной формации.
6)Наличие представительства всех функций нижележащих структур ЦНС.
7)Цитоархитектоническое распределение на поля.
8)Наличие в специфических проекционных сенсорных и моторных системах коры вторичных и третичных полей с преобладанием ассоциативных функций.
9)Наличие специализированных ассоциативных областей коры (центров).
10)Динамическая локализация функций, выражающаяся в возможности компенсаций функций утраченных структур коры.
11)Перекрытие в коре зон соседних периферических рецептивных полей.
12)Возможность длительного сохранения следов раздражения.
13)Реципрокная функциональная взаимосвязь возбудительных и тормозных состояний коры.
14)Способность к иррадиации состояний возбуждения и торможения.
15)Наличие специфической электрической активности
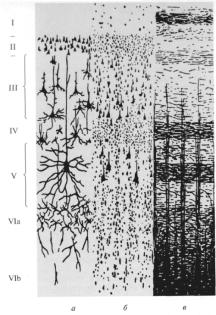
Вкоре выделяют 6 слоев (I–VI), каждый из которых состоит из пирамидных и звездчатых клеток (рис. 1). Слои коры больших полушарий:
1)Молекулярный представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, среди которых расположены редкие горизонтальные клетки и клетки-зерна, сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры большого мозга.
2)Наружный зернистый слой состоит из звездчатых клеток, определяющих длительность циркуляции возбуждения в коре большого мозга, т. е. имеющих отношение к памяти.
3) |
Наружный |
пирамидный |
слой |
формируется |
из пирамидных |
клеток |
|
|
малой величины и функционально вместе со 2 слоем обеспечивает корково-корковые |
||||||
|
связи различных извилин мозга. |
|
|
|
|
|
|
4) |
Внутренний |
зернистый |
слой |
содержит |
звездчатые |
клетки, |
здесь |
|
заканчиваются специфические таламокортикальные пути, т. е. пути, начинающиеся от |
||||||
|
рецепторов анализаторов. |
|
|
|
|
|
|
CerebruM |
1 |
подготовил Дадаев Гаджимурад
5)Внутренний пирамидный слой крупных нейронов, имеющих форму пирамид, являются выходными нейронами, их аксоны идут в продолговатый и спинной мозг.
6)Слой полиморфных клеток. Большинство нейронов этого слоя образуют кортикоталамические пути.
Рис. 1. Полусхематичное изображение слоев коры головного мозга (по Бродману):
а – основные типы нервных клеток (окраска по Гольджи); б – тела нейронов (окраска по Нисслю); в – общее расположение волокон (миелиновых оболочек).
Главная особенность пирамидных клеток заключается в том, что их аксоны выходят из коры и оканчиваются в других корковых или иных структурах. Название звездчатых клеток также обусловлено их формой; их аксоны оканчиваются в коре, т.е. речь идет о корковых интернейронах.
ВI – IV слоях происходит восприятие и обработка поступающих в кору сигналов в виде нервных импульсов. Покидающие кору эфферентные пути формируются преимущественно в V– VI слоях. Деление коры на различные поля проведено К. Бродманом (1909) на основе цитоархитектонических признаков – формы и расположения нейронов.
Нейронный состав, его распределение по слоям различно в разных областях коры, что позволило выделить в мозге человека 53 цитоархитектонических поля. Эти поля в определенной степени соответствуют функциональным и нейрохимическим зонам коры. Первичные поля имеют рядом расположенные вторичные и третичные поля, обеспечивающие взаимодействие функций данного анализатора с функциями других анализаторов. Для всех анализаторов характерен соматотопический принцип организации проекции на кору периферических рецепторных систем.
1.1.Локализация функций в коре большого мозга
Внастоящее время принято подразделение коры на сенсорные, двигательные и ассоциативные (неспецифические) зоны (области).
Сенсорные зоны коры. Сенсорная информация поступает в проекционную кору, корковые отделы анализаторов (И.П. Павлов). Эти зоны расположены преимущественно в теменной, височной и затылочной долях. Восходящие пути в сенсорную кору поступают в основном от релейных сенсорных ядер таламуса.
Первичные сенсорные зоны – это зоны сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения чувствительности организма (ядра анализаторов по И. П. Павлову). Они состоят из мономодальных нейронов и формируют
CerebruM |
2 |

подготовил Дадаев Гаджимурад
ощущения одного качества. В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, их рецепторных полей.
Первичные проекционные зоны коры состоят главным образом из нейронов 4-го афферентного слоя, для которых характерна четкая топическая организация. Значительная часть этих нейронов обладает высочайшей специфичностью. Так, например, нейроны зрительных областей избирательно реагируют на определенные признаки зрительных раздражителей: одни – на оттенки цвета, другие – на направление движения, третьи – на характер линий (край, полоса, наклон линии) и т.п. Однако следует отметить, что в первичные зоны отдельных областей коры включены также нейроны мультимодального типа, реагирующие на несколько видов раздражителей. Кроме того, там же имеются нейроны, реакция которых отражает воздействие неспецифических (лимбико-ретикулярных, или модулирующих) систем.
Вторичные сенсорные зоны расположены вокруг первичных сенсорных зон, менее локализованы, их нейроны отвечают на действие нескольких раздражителей, т.е. они полимодальны.
Локализация сенсорных зон. Важнейшей сенсорной областью является теменная доля постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий. Эту зону обозначают как соматосенсорную область I. Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности и чувствительности опорнодвигательного аппарата – от мышечных, суставных, сухожильных рецепторов (рис. 2).
Рис. 2. Схема чувствительного и двигательного гомункулусов (по У. Пенфильду, Т. Расмуссену). Разрез полушарий во фронтальной плоскости:
а – проекция общей чувствительности в коре постцентральной извилины; б – проекция двигательной системы в коре предцентральной извилины
Кроме соматосенсорной области I выделяют соматосенсорную область II меньших размеров, расположенную на границе пересечения центральной борозды с верхним краем височной доли, в глубине латеральной борозды. Точность локализации частей тела здесь выражена в меньшей степени. Хорошо изученной первичной проекционной зоной является слуховая кора (поля 41, 42), которая расположена в глубине латеральной борозды (кора поперечных височных извилин Гешля). К проекционной коре височной доли относится также центр вестибулярного анализатора в верхней и средней височных извилинах.
В затылочной доле расположена первичная зрительная область (кора части клиновидной
CerebruM |
3 |
подготовил Дадаев Гаджимурад
извилины и язычковой дольки, поле 17). Здесь имеется топическое представительство рецепторов сетчатки. Каждой точке сетчатки соответствует свой участок зрительной коры, при этом зона желтого пятна имеет сравнительно большую зону представительства. В связи с неполным перекрестом зрительных путей в зрительную область каждого полушария проецируются одноименные половины сетчатки. Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения. Около поля 17 расположена кора вторичной зрительной области (поля 18 и 19). Нейроны этих зон полимодальны и отвечают не только на световые, но и на тактильные и слуховые раздражители. В данной зрительной области происходит синтез различных видов чувствительности, возникают более сложные зрительные образы и их опознание.
Во вторичных зонах ведущими являются 2-й и 3-й слои нейронов, для которых основная часть информации об окружающей среде и внутренней среде организма, поступившая в сенсорную кору, передается для дальнейшей ее обработки в ассоциативную кору, после чего инициируется (в случае необходимости) поведенческая реакция с обязательным участием двигательной коры.
Двигательные зоны коры. Выделяют первичную и вторичную моторные зоны.
В первичной моторной зоне (прецентральная извилина, поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела (см. рис. 2). Основной закономерностью топографического представительства является то, что регуляция деятельности мышц, обеспечивающих наиболее точные и разнообразные движения (речь, письмо, мимика), требует участия больших по площади участков двигательной коры. Раздражение первичной моторной коры вызывает сокращение мышц противоположной стороны тела (для мышц головы сокращение может быть билатеральное). При поражении этой корковой зоны утрачивается способность к тонким координированным движениям конечностями, особенно пальцами рук.
Вторичная моторная зона (поле 6) расположена как на латеральной поверхности полушарий, впереди прецентральной извилины (премоторная кора), так и на медиальной поверхности, соответствующей коре верхней лобной извилины (дополнительная моторная область). Вторичная двигательная кора в функциональном плане имеет главенствующее значение по отношению к первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием и координацией произвольных движений. Здесь в наибольшей степени регистрируется медленно нарастающий отрицательный потенциал готовности, возникающий примерно за 1 с до начала движения. Кора поля 6 получает основную часть импульсации от базальных ганглиев и мозжечка, участвует в перекодировании информации о плане сложных движений.
Раздражение коры поля 6 вызывает сложные координированные движения, например, поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения сгибателей или разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины (поле 6), центр моторной речи Брока в заднем отделе нижней лобной извилины (поле 44), обеспечивающие речевой праксис, а также музыкальный моторный центр (поле 45), обеспечивающий тональность речи, способность петь. Нейроны двигательной коры получают афферентные входы через таламус от мышечных, суставных и кожных рецепторов, от базальных ганглиев и мозжечка. Основным эфферентным выходом двигательной коры на стволовые и спинальные моторные центры являются пирамидные клетки V слоя. Основные доли коры большого мозга представлены на рис. 3.
CerebruM |
4 |

подготовил Дадаев Гаджимурад
Рис. 3. Четыре основные доли коры головного мозга (лобная, височная, теменная и затылочная); вид сбоку. В них расположены первичная двигательная и сенсорная области, двигательные и сенсорные
области более высокого порядка (второго, третьего и т.д.) и ассоциативная (неспецифичная) кора
Ассоциативные области коры (неспецифическая, межсенсорная, межанализаторная кора) включают участки новой коры большого мозга, которые расположены вокруг проекционных зон
ирядом с двигательными зонами, но не выполняют непосредственно чувствительных или двигательных функций, поэтому им нельзя приписывать преимущественно сенсорные или двигательные функции, нейроны этих зон обладают большими способностями к обучению. Границы этих областей обозначены недостаточно четко. Ассоциативная кора является филогенетически наиболее молодой частью новой коры, получившей наибольшее развитие у приматов и у человека. У человека она составляет около 50% всей коры или 70 % неокортекса. Термин «ассоциативная кора» возник в связи с существовавшим представлением о том, что эти зоны за счет проходящих через них кортико-кортикальных соединений связывают двигательные зоны и одновременно служат субстратом высших психических функций. Основными
ассоциативными зонами коры являются: теменно-височно-затылочная, префронтальная кора лобных долей и лимбическая ассоциативная зона.
Нейроны ассоциативной коры являются полисенсорными (полимодальными): они отвечают, как правило, не на один (как нейроны первичных сенсорных зон), а на несколько раздражителей, т. е. один и тот же нейрон может возбуждаться при раздражении слуховых, зрительных, кожных и др. рецепторов. Полисенсорность нейронов ассоциативной коры создается кортико-кортикальными связями с разными проекционными зонами, связями с ассоциативными ядрами таламуса. В результате этого ассоциативная кора представляет собой своеобразный коллектор различных сенсорных возбуждений и участвует в интеграции сенсорной информации
ив обеспечении взаимодействия сенсорных и моторных областей коры.
Ассоциативные области занимают 2-й и 3-й клеточные слои ассоциативной коры, на которых происходит встреча мощных одномодальных, разномодальных и неспецифических афферентных потоков. Работа этих отделов коры мозга необходима не только для успешного синтеза и дифференцировки (избирательного различения) воспринимаемых человеком раздражителей, но и для перехода к уровню их символизации, т. е. для оперирования значениями слов и использования их для отвлеченного мышления, для синтетического характера восприятия.
В настоящее время по таламокортикальным проекциям предлагают выделять три основные а с с о ц и а т и в н ы е с и с т е м ы м о з г а : таламотеменную, таламолобную и таламовисочную.
CerebruM |
5 |
подготовил Дадаев Гаджимурад
Таламотеменная система представлена ассоциативными зонами теменной коры, получающими основные афферентные входы от задней группы ассоциативных ядер таламуса. Теменная ассоциативная кора имеет эфферентные выходы на ядра таламуса и гипоталамуса, в моторную кору и ядра экстрапирамидной системы. Основными функциями таламотеменной системы являются гнозис и праксис. Под гнозисом понимают функцию различных видов узнавания: формы, величины, значения предметов, понимание речи, познание процессов, закономерностей и др. К гностическим функциям относится оценка пространственных отношений, например, взаимного расположения предметов. В теменной коре выделяют центр стереогнозиса, обеспечивающий способность узнавания предметов на ощупь. Вариантом гностической функции является формирование в сознании трехмерной модели тела («схемы тела»). Под праксисом понимают целенаправленное действие. Центр праксиса находится в надкорковой извилине левого полушария, он обеспечивает хранение и реализацию программы двигательных автоматизированных актов.
Таламолобная система представлена ассоциативными зонами лобной коры, имеющими основной афферентный вход от ассоциативного медиодорсального ядра таламуса, других подкорковых ядер. Основная роль лобной ассоциативной коры сводится к инициации базовых системных механизмов формирования функциональных систем целенаправленных поведенческих актов (П. К.Анохин). Префронтальная область играет главную роль в выработке стратегии поведения. Нарушение этой функции особенно заметно, когда необходимо быстро изменить действие и когда между постановкой задачи и началом ее решения проходит некоторое время, т.е. успевают накопиться раздражители, требующие правильного включения в целостную поведенческую реакцию.
Таламовисочная система. Некоторые ассоциативные центры, например, стереогнозиса, праксиса, включают в себя и участки височной коры. В височной коре расположен слуховой центр речи Вернике, находящийся в задних отделах верхней височной извилины левого полушария. Этот центр обеспечивает речевой гнозис: распознание и хранение устной речи как собственной, так и чужой. В средней части верхней височной извилины находится центр распознания музыкальных звуков и их сочетаний. На границе височной, теменной и затылочной долей находится центр чтения, обеспечивающий распознание и хранение образов.
Существенную роль в формировании поведенческих актов играет биологическое качество безусловной реакции, а именно ее значение для сохранения жизни. В процессе эволюции — это значение было закреплено в двух противоположных эмоциональных состояниях – положительном и отрицательном, которые у человека составляют основу его субъективных переживаний - удовольствия и неудовольствия, радости и печали. Во всех случаях целенаправленное поведение строится в соответствии с эмоциональным состоянием, возникшим при действии раздражителя. Во время поведенческих реакций отрицательного характера напряжение вегетативных компонентов, особенно сердечно-сосудистой системы, в отдельных случаях, особенно в непрерывных так называемых конфликтных ситуациях, может достигать большой силы, что вызывает нарушение их регуляторных механизмов (вегетативные неврозы).
1.2. ЭЭГ. Корковые ритмы.
Электроэнцефалография - регистрация суммарной электрической активности мозга с поверхности головы, а электроэнцефалограмма (ЭЭГ) представляет собой кривую, зарегистрированную при этом. Запись ЭЭГ с коры головного мозга называется
электрокортикограммой (ЭКоГ).
Регистрация ЭЭГ производится с помощью биполярных (оба активны) или униполярных (активный и индифферентный) электродов, накладываемых симметрично в лобных, центральных, теменных, височных и затылочных областях головного мозга. Основными анализируемыми параметрами ЭЭГ являются частота и амплитуда волновой активности.
На ЭЭГ (рис. 4) регистрируется 4 основных физиологических ритма: -, -, - и - ритмы (альфа, бета, гамма, дельта).
CerebruM |
6 |

подготовил Дадаев Гаджимурад
Рис. 4.1. Электроэнцефалограмма:
а – основные ритмы: 1 – -ритм; 2 – -ритм; 3 – -ритм; 4 – -ритм; б – реакция десинхронизации
ЭЭГ затылочной области коры при открывании глаз (↑) и восстановление-ритма при закрывании глаз (↓)
-Ритм имеет частоту 8–13 Гц, амплитуду до 70 мкВ, наблюдается у человека в состоянии физического, интеллектуального и эмоционального покоя. -Ритм является упорядоченным регулярным ритмом. Если он доминирует, ЭЭГ рассматривается как синхронизированная. Механизм синхронизации ЭЭГ связан с деятельностью выходных ядер таламуса (Экклс).
-Ритм преобладает у 85 –95% здоровых людей старше девятилетнего возраста. Лучше всего он выражен в затылочных областях мозга, в передних (центральной и лобной) областях, часто сочетается с -ритмом. Вариантом -ритма являются «веретена сна» длительностью 2 –8 с, которые наблюдаются при засыпании и представляют собой регулярные чередования нарастания
иснижения амплитуды волн в частотах -ритма.
-Ритм имеет нерегулярную частоту 14 – 30 Гц, низкую амплитуду – до 30 мкВ, сменяет-ритм при сенсорной стимуляции, например, при действии света, при эмоциональном возбуждении. Наиболее выражен -ритм в лобных, центральных областях головного мозга. - Ритм отражает высокий уровень функциональной активности головного мозга. Смена -ритма - ритмом называется десинхронизацией ЭЭГ и объясняется активирующим влиянием на кору больших полушарий восходящей ретикулярной формации ствола и лимбической системы.
-Ритм имеет частоту 4 – 7 Гц, амплитуду до 200 мкВ. У бодрствующего человека θ-ритм регистрируется обычно в передних областях мозга при длительном эмоциональном напряжении и почти всегда регистрируется в процессе развития фаз медленноволнового сна. Отчетливо регистрируется у детей, пребывающих в состоянии неудовольствия.
-Ритм имеет частоту 0,5 – 3,0 Гц, амплитуду 200 – 300 мкВ. Эпизодически регистрируется во всех областях головного мозга. Появление этого ритма у бодрствующего человека свидетельствует о снижении функциональной активности мозга. Стабильно фиксируется во время глубокого медленноволнового сна.
Происхождение - и -ритмов ЭЭГ связывают с активностью соответственно мостовой и бульбарной синхронизирующих систем ствола мозга.
CerebruM |
7 |
подготовил Дадаев Гаджимурад
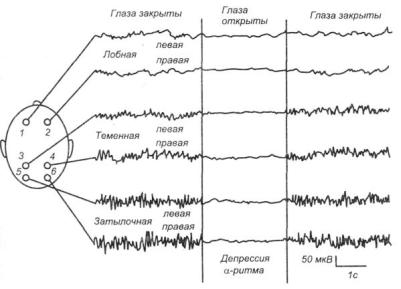
Рис. 5. ЭЭГ бодрствующего человека при шести отведениях с поверхности головы
Показатели ЭЭГ. При малейшем привлечении внимания к любому стимулу развивается десинхронизация ЭЭГ - реакция блокады -ритма (рис. 5). Хорошо выраженный -ритм – показатель покоя организма (релаксации). Более сильная реакция активации выражается не только в блокаде -ритма, но и в усилении высокочастотных составляющих ЭЭГ'. - и -
активности. Падение уровня функционального состояния выражается в уменьшении доли высокочастотных составляющих и росте амплитуды у более медленных ритмов – - и - колебаний. Частотные спектры ЭЭГ помогают количественно оценивать динамику ее изменения. Выделяют несколько типов частотных спектров фоновой ЭЭГ бодрствования: ЭЭГ с -ритмом (1) и без -ритма (2), а также с преобладанием -активности (3) и спектра ЭЭГ десинхронизированного типа, когда ни один из ритмов не доминирует (4).
2. Понятие о высшей и низшей нервной деятельности
Строение нервной системы и совокупность процессов, проходящих в ней, позволяют выполнять регулирующие и управляющие функции, которые обеспечивают:
1)быстрое координирование функций организма;
2)согласование состояния организма с различными условиями окружающей среды;
3)объединение отдельных органов и систем организма в единое целое.
Аппарат управления у высших животных и человека представлен рефлекторным механизмом, который проявляется во всех отделах нервной системы и является основной формой деятельности нервной системы.
В настоящее время рефлексом называется всякая реакция организма, возникающая при действии раздражителя из внешней или внутренней среды и осуществляемая при обязательном участии ЦНС.
Впервые представление о рефлекторном принципе работы высших отделов ЦНС было выдвинуто и обосновано И.М.Сеченовым в книге «Рефлексы головного мозга» (1863). Идеи И.М.Сеченова были развиты И.П.Павловым. В период с 1901 по 1936 гг. им было создано учение о высшей нервной деятельности. Все разнообразие рефлекторных актов И. П. Павлов разделил на два вида рефлексов: безусловные и условные рефлексы.
Под безусловными рефлексами он понимал врожденные, постоянные, видоспецифические реакции организма, носящие наследственный характер. Совокупность безусловных рефлексов, по Павлову, составляет основу низшей нервной деятельности (деятельности спинного мозга, ствола головного мозга и мозжечка).
CerebruM |
8 |

подготовил Дадаев Гаджимурад
Условными рефлексами И. П. Павлов назвал индивидуальные, временные реакции организма, вырабатываемые на основе жизненного опыта и не передающиеся по наследству. Совокупность условных рефлексов составляет основу высшей нервной деятельности – деятельности коры больших полушарий и ближайших подкорковых структур.
Согласно учению И.П. Павлова, активное взаимодействие организма и среды осуществляется благодаря низшей и высшей нервной деятельности.
Низшая нервная деятельность (ННД) – это совокупность нейрофизиологических процессов, обеспечивающих осуществление безусловных рефлексов и инстинктов.
ННД обеспечивает регуляцию деятельности внутренних органов и их взаимосвязь, благодаря чему организм функционирует как единое целое.
Инстинкт (лат. instinctus — побуждение) — это врожденная приспособительная стереотипная деятельность организма, побуждаемая основными биологическими потребностями и внешними раздражителями. Инстинкты осуществляются с участием структур промежуточного и конечного мозга.
Высшая нервная деятельность (ВНД) - это совокупность нейрофизиологических процессов, обеспечивающих сознательную и подсознательную переработку и усвоение информации, индивидуальное приспособительное поведение организма в окружающей среде, в том числе трудовая деятельность, обучение в онтогенезе всем видам деятельности, в том числе целенаправленному поведению в обществе.
ВНД обеспечивает взаимосвязь организма с окружающей средой. ВНД – это аналитикосинтетическая деятельность коры и ближайших подкорковых образований, которая проявляется в способности выделять из окружающей среды ее отдельные элементы и объединять их в комбинации.
Таким образом, с помощью ВНД осуществляется психическая деятельность. Психическая деятельность – это идеальная субъективно осознаваемая деятельность организма, осуществляемая с помощью нейрофизиологических процессов.
Психическая деятельность протекает только в период бодрствования и осознается, а ВНД – и в период сна как неосознаваемая переработка информации, и в период бодрствования как осознаваемая и подсознательная переработка.
3. Отличия условных рефлексов от безусловных рефлексов
Безусловные рефлексы |
Условные рефлексы |
|
|
||
|
|
|
Врожденные, отражают видовые особенности |
Приобретаются в течение жизни, отражают |
|
организма |
индивидуальные особенности организма |
|
|
|
|
Относительно постоянны в течение жизни особи |
Образуются, изменяются и отменяются, когда они |
|
|
становятся неадекватными условиям жизни особи |
|
|
|
|
Реализуются по анатомическим путям, |
Реализуются по функционально-организующимся |
|
определенным генетически |
временным (замыкательным) связям |
|
|
|
|
Свойственны всем уровням ЦНС и |
Для своего образования и реализации требуют |
|
осуществляются преимущественно ее низшими |
целостности коры большого мозга, особенно у |
|
отделами (спинной мозг, стволовой отдел, |
высших млекопитающих животных |
|
подкорковые ядра) |
|
|
|
|
|
Каждый рефлекс имеет свое специфическое |
Рефлексы могут образовываться с любого |
|
рецептивное поле и специфические раздражители |
рецептивного поля на самые разнообразные |
|
|
раздражители |
|
|
|
|
|
|
|
Реагируют на действие наличного раздражителя, |
Приспосабливают организм к действию стимула, |
|
которого уже нельзя избежать |
которое еще предстоит испытать, т.е. имеют |
|
|
предупредительное значение |
|
|
|
|
CerebruM |
9 |
подготовил Дадаев Гаджимурад
4. Классификация условных рефлексов
Условные рефлексы подразделяют по нескольким критериям.
1.По биологическому значению различают:
пищевые;
половые;
оборонительные;
двигательные;
ориентировочный - реакция на новый раздражитель.
Ориентировочный рефлекс осуществляется в 2 фазы:
1)стадия неспецифической тревоги - 1-я реакция на новый раздражитель: изменяются двигательные реакции, вегетативные реакции, изменяется ритм ЭЭГ; продолжительность этой стадии зависит от силы и значимости раздражителя.
2)стадия исследовательского поведения: восстанавливается двигательная активность, вегетативные реакции, ритм ЭЭГ; возбуждение охватывает большой отдел коры головного мозга и образования лимбической системы; результат - познавательная деятельность.
Отличия ориентировочного рефлекса от других условных рефлексов:
врожденная реакция организма;
он может угасать при повторении действия раздражителя.
То есть ориентировочный рефлекс занимает промежуточное место между безусловным и условным рефлексом.
2.По характеру афферентного звена (по виду рецепторов), с которых идет выработка, условные рефлексы делят:
экстерорецептивные - формируют приспособительное поведение животных по добыванию пищи, избеганию вредных воздействий, продолжению рода и т.д. Для человека важнейшее значение имеют экстерорецептивные словесные раздражители, формирующие поступки и мысли;
проприорецептивные – лежат в основе научения животных и человека двигательным навыкам: ходьбе, производственным операциям и др.;
интерорецептивные – влияют на настроение, работоспособность.
3.По характеру эфферентного ответа (по отделу нервной системы) различают:
соматические (двигательные);
вегетативные (сердечно-сосудистые, секреторные, выделительные, спинальные, бульбарные, кортикальные).
4.В зависимости от сигнальной системы различают
УР на сигналы первой сигнальной системы (органы чувств)
УР на сигналы второй сигнальной систем (на речь), вырабатываются только у человека.
5.По особенностям подкрепления:
УР первого порядка - если в качестве подкрепления используется безусловный раздражитель, эти рефлексы также вырабатываются первыми в процессе онтогенеза. Пример: если ребенка в возрасте 3-4 месяца кормить только из красной бутылочки, он безошибочно выберет ее среди других: условный раздражитель – бутылочка, подкрепляющий – пища.
УР второго порядка - если в качестве подкрепления используется раздражитель, вызывающий ранее выработанный прочный условный рефлекс.
Пример: если перед тем, как дать ребенку в 6-8 месяцев пищу в красной бутылочке,
CerebruM |
10 |