

2 курс / Нормальная физиология / Молекулярная_физиология_Пятин_В_Ф_,_Баишева_Г_М_и_др_
.pdfПРИЛОЖЕНИЕ 23 |
|
ПРИЛОЖЕНИЕ 24 |
ПРИЛОЖЕНИЕ 23. МЕХАНИЗМ ПЕРЕДАЧИ ВОЗБУЖДЕНИЯ В |
ПРИЛОЖЕНИЕ 24. СТРОЕНИЕ И ЖИЗНЕННЫЙ ЦИКЛ |
|
ЭЛЕКТРИЧЕСКОМ И ХИМИЧЕСКОМ СИНАПСАХ |
СИНАПТИЧЕСКИХ ВЕЗИКУЛ |
|
А |
Б |
А. У электрического синапса клеточные мембраны соседних нейронов тесно прилегают друг к другу, так что между ними остается только очень узкая щель шириной 2 нм. В зоне сближения мембран, называемой щелевым контактом (gap junction), в каждой из них имеются специфические белковые комплексы. Они состоят из шести субъединиц (коннексинов) и располагаются в таком порядке, что в их центре образуется пора, заполненная водой, которая проходит через бислой клеточной мембраны. Эти протеиновые комплексы, названные коннексонами, располагаются напротив друг друга так, что поры одной клетки образуют с порами другой клетки открытые связи, т.е. возникают «каналы». Основное функциональноезначение:быстрововлечьвпроцессвозбуждениябольшоечислоклеток(функциональный синцитий).
Б. Химический синапс имеет в пресинаптической мембране везикулы, заполненные трансмиттером (медиатором), широкую синаптическую щель (30-50 нм), на постсинаптической мембране расположены рецепторы к трансмиттерам. Молекулы медиатора, высвобождаемые из пресинапса, связываются с рецепторами на постсинаптической мембране, в результате чего прямо или опосредованно открываются ионные каналы. Ионы, которые начинают поступать внутрь постсинаптической клетки через открытые каналы, изменяют заряд её мембраны, что является частичной гиперполяризацией (в случае тормозного синапса) или деполяризацией (в случае возбуждающего синапса) этой мембраны и, как следствие, приводит к торможению или провоцированию генерации постсинаптической клеткой потенциала действия.
Жизненнный цикл синаптической везикулы. В аппарате Гольджи сомы ней-
рона формируются мембранные образования в виде пузырьков, не заполненных медиатором. Эти пузырьки направляются в пресинаптическое окончание с помощью системы быстрого аксонного транспорта. В пресинаптическом окончании пузырьки заполняются медиаторами посредством АТФ-зависимого протонного насоса. Молекулы протонной АТФазы входят в состав мембраны синаптических пузырьковиподдерживаютопределенныйуровеньмембранногопотенциала.Мембрана синаптического пузырька содержит также стимулируемую кальмодулином Са2+-АТФазу, которая обеспечивает поглощение пузырьками ионов Са2+. Популяция зрелых (заполненных) пузырьков подвергается экзоцитозу, освобождая содержимое в синаптическую щель. Освобождение медиатора может осуществляться посредствомэкзоцитоза сполным («классический» механизм) либо неполным (механизм «kiss-and-run fusion») слиянием. Первый вид экзоцитоза сопровождается встраиванием мембраны везикулы в пресинаптическую, опорожнением пузырька, а затем посредством эндоцитоза образуются покрытые клатрином везикулы, которые затем проходят стадию эндосомы и снова заполняются медиатором. Второй вид экзоцитоза характеризуется образованием временной поры, соединяющей полость пузырька с синаптической щелью. После выделения медиатора везикула не встраивается в пресинаптическую мембрану, а отпочковывается от неё и повторно заполняется медиатором. Циклы экзоцитоз — эндоцитоз повторяются.
140 |
141 |

ПРИЛОЖЕНИЕ 25
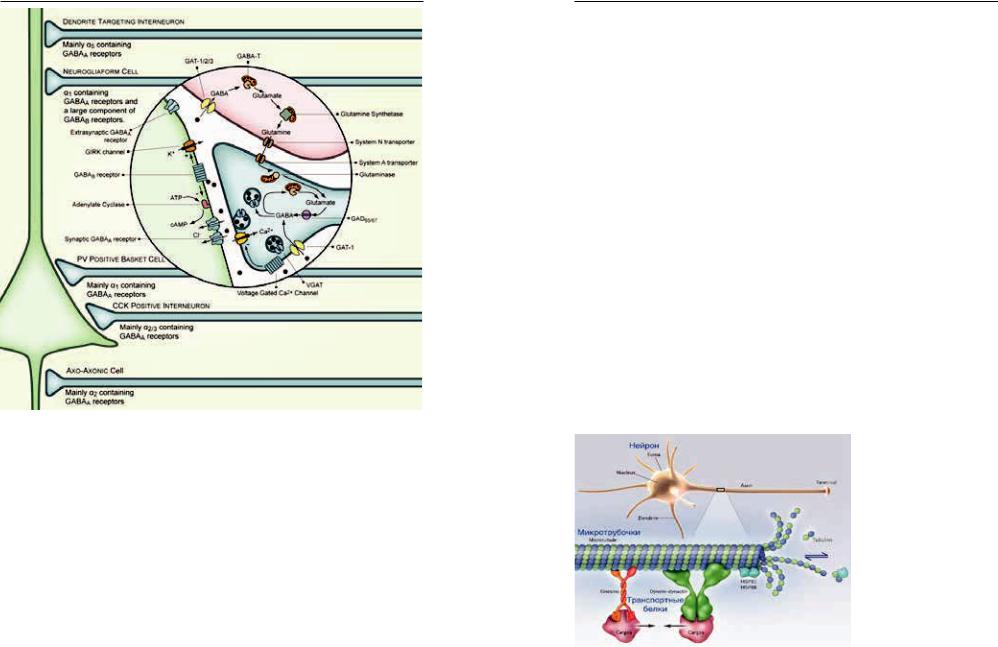
ПРИЛОЖЕНИЕ 25. СХЕМА ГАМК-ЕРГИЧЕСКОЙ ПЕРЕДАЧИ
Эффект ГАМК заключается в гиперполяризации мембраны и снижении вероятности прохождения ПД даже в случае одновременного получения клеткой возбуждающего сигнала. Таким образом достигается баланс между возбуждающими и тормозными сигналами, приходящими к клетке с интервалами примерно в 10 мс, который и обусловливает ее текущую активность.
Для производства ГАМК используется глутамин и его производные. Транспортер — белковая молекула, транспортирующая молекулы медиатора обратно в клетку для повторного использования. Ауторецепторы принимают участие в саморегуляции синапса. GAT– переносчик ГАМК, GAD – глутаматдекарбоксилаза, VIAAT – везикулярный транспортер тормозных медиаторов, Gin – глутамин, Glu – глутаминовая кислота.
Известно большое количество подтипов ГАМК рецепторов. Так, наиболее известные ГАМК-А рецепторы чувствительны к бензодиазепинам, бициклофосфатам, барбитуратам, пикротоксину.
Некоторые ГАМК рецепторы ассоциированы с калиевыми, кальциевыми каналами и G-белком: они модулируют высвобождение катехоламинов и других нейромедиаторов в головном мозге.
Взаимодействие ГАМК с рецепторами постсинаптической мембраны приводит к конформационным изменениям хлор-ионного канала. При этом для открытия канала требуется присоединение не менее двух молекул ГАМК. Усиление тока хлора в клетку приводит к понижению возбудимости нейронов.
ПРИЛОЖЕНИЕ 26
ПРИЛОЖЕНИЕ 26. СХЕМА ГЛУТАМАТЕРГИЧЕСКОЙ ПЕРЕДАЧИ
Девяностопроцентовнейроновкорыиспользуютглутамат –основной возбуждающий медиатор, увеличивающий вероятность развития аксонного потенциала действия на постсинаптическом нейроне при высвобождении в синаптическую щель. В человеческом мозге глутамат чаще всего используется крупными пирамидными нейронами коры и более глубоких структур мозга. Также этот медиатор нередко используется в модифицируемых синапсах, обусловливая обучение. Поскольку глутамат представляет собой важнейший трансмиттер ЦНС, в качестве средства для наркоза применяют фармакологический препарат кетамин, представляющий собой антагонист глутамата.
Глутаматергическая сигнализация требует участия трех клеток. Глюкоза является одним из промежуточных метаболитов при синтезе глутамата. Глутамат прямо открывает неспецифический ионный канал для катионов. Существует, по крайней мере, три основных типа постсинаптических рецепторов, у каждого из которых много подтипов. Они различаются по своей способности связываться с экзогенными агонистами. К этим рецепторам относятся ионотропные рецепторы - лиганд- управляемыекатионныеканалы-NMDA(N-метил-D-аспартат),AMPA(α-амино-3- гидрокси-5-метил-4-изоксазол-пропионовая кислота) и метаботропный mGluR1-5, действие которого осуществляется через ИФ3/ДАГ (инозитолтрифосфат/диацилглицерол).
Через катионный канал AMPA в постсинаптическую область входят ионы Na+, которыедеполяризуютпостсинаптическуюмембрану.ВотличиеотAMPA,NMDA-
142 |
143 |

ПРИЛОЖЕНИЕ 25
каналоткрывается,еслипостсинаптическаямембранадостаточнодеполяризована. Через этот канал в постсинаптическую область поступают и ионы Na+, и Са2+. Вход Са2+ в клетку приводит к активации протеинкиназы С. Параллельно Са2+ связывается в комплекс с кальмодулином и активирует Са2+/кальмодулин киназу II. Оба этих процесса направлены на фосфорилирование синаптических белков. В итоге запускается процесс введения дополнительныхAMPAрецепторов в постсинапти- ческуюмембрану,которыйспособствуетдлительномупотенцированию-долговре-
менному синаптическому потенцированию (Long-Term potentiation - LTP).
Стоит отметить, что чрезмерное возбуждение многих NMDA синапсов может также необратимо повредить постсинаптические клетки (так называемая эксайтотоксичность - цитотоксичность, свойственная возбуждающим нейротрансмиттерам). Эксайтотоксичность усиливает многие неврологические заболевания, такие, как нарушение слуха, течение болезни Альцгеймера или наследственные повреждения, вызванные первичной гипоксией.
После реализации возбуждающего эффекта глутамат из синаптической щели удаляется либо посредством обратного захвата в пресинаптическую область, либо в клетки глии.
ПРИЛОЖЕНИЕ 27
ПРИЛОЖЕНИЕ 27. СХЕМА ХОЛИНЕРГИЧЕСКОЙ ПЕРЕДАЧИ
Среди синапсов с химическим механизмом передачи электрического сигнала особое место занимает холинергический синапс, у которого медиатором служит ацетилхолин (АЦХ). Особенность этих синапсов заключается в том, что в организме присутствуют две их группы, подразделяющиеся по тому типу АЦХ-рецептора, который присутствует на постсинаптической мембране. Это АЦХ-рецепторы, чувствительные дополнительно к никотину (нАЦХ-рецептор и, соответственно, нАЦХ синапс), и АЦХ-рецепторы, чувствительные дополнительно к мускарину (мАЦХ-рецептор и, соответственно, мАЦХ синапс). АЦХ синапсы с никотиновым АЦХ-рецепторомявляютсяионотропными,аАЦХсинапсысмускариновымАЦХ- рецептором - метаботропными.
В ионотропном никотиновом АЦХ синапсе две молекулы свободного АЦХ связываютсясрецепторнымбелком,представляющимсобойоднуизструктурионного канала. Активация нАЦХ-канала вызывает проводимость постсинаптической мембраныдлякатионов,деполяризациюпостсинаптическоймембраны,формирование потенциала и, в итоге, сокращение мышцы. В метаботропном мускариновом АЦХ синапсе рецептором служит белок, который не является ионным каналом. Ионные каналы, необходимые для возникновения потенциала на постсинаптической мембране, открываются благодаря вторичным посредникам. Как только трансмиттер связывается с рецептором, G-белок, имеющий три субъединицы, образует с рецептором комплекс. ГДФ, связанный с G-белком, заменяется на ГТФ. При этом образуется активированный G-белок, состоящий из ГТФ и α-субъединицы, который открывает калиевый ионный канал.
144 |
145 |

ПРИЛОЖЕНИЕ 28
ПРИЛОЖЕНИЕ 28. СХЕМА АДРЕНЕРГИЧЕСКОЙ ПЕРЕДАЧИ
Норадреналин (норэпинефрин - НА или НЭ) накапливается в пузырьках (везикулах; диаметр 0,05-0,2 мкм), окруженных мембраной. Норадреналин образуется в результате последовательного ферментативного синтеза из аминокислоты тирозина, которая при участии фермента тирозингидроксилазы превращается в L-допа. L-допадекарбоксилируетсясобразованиемдопамина,которыйспомощьювезику- лярного транспорта моноаминов (ВМАТ) попадает в везикулы. И только в везикулах под действием допамин-β-гидроксилазы допамин превращается в норадреналин. В надпочечниках большая часть норадреналина превращается в адреналин в результате протекающего в цитозоле в одну стадию ферментативного метилирования. При электрическом возбуждении симпатического нерва содержимое везикулы, в том числе норадреналин, высвобождается в синаптическую щель.
В ответ на нервные импульсы происходит высвобождение норадреналина в синаптическую щель и последующее его взаимодействие с адренорецепторами постсинаптической, а также пресинаптической мембраны. Существующие в организме адренорецепторы обладают неодинаковой чувствительностью к химическим соединениям. Так, выделяют α- и β-адренорецепторы. α1-подтип адренорецепторы локализуются постсинаптически, α2-адренорецепторы - пресинаптически и вне синапсов. Физиологическая роль пресинаптических α2-адренорецепторов заключается в их участии в системе обратной отрицательной связи, регулирующей высвобождение норадреналина. Возбуждение этих рецепторов норадреналином (или другими веществами, обладающими α2-адреномиметической активностью) тормозит высвобождение норадреналина из варикозных утолщений. α2-Адренорецепторы расположены и на мембранах эффекторных клеток, вне синапсов. Среди β-адренорецепторов выделяют β1-адренорецепторы (например, в
ПРИЛОЖЕНИЕ278
сердце), β2-адренорецепторы (в бронхах, сосудах, матке) и β3-адренорецепторы (в жировой ткани). Обнаружены также пресинаптические β-адренорецепторы (β2подтип): в отличие от аналогичных α-адренорецепторов они осуществляют положительную обратную связь, стимулируя высвобождение норадреналина.
Действие свободного норадреналина быстро прекращается. Почти 90% норадреналина быстро захватывается специфическим транспортным механизмом (NAT) из синаптической щели в аксоплазму и вновь заключается в везикулы по механизму везикулярного транспорта (обратный нейрональный захват – re-uptake медиатора). Транспорт норадреналина (NAT) может тормозиться трициклическими антидепрессантами и кокаином. Кроме того, норадреналин из синаптической щели доставляется к эффекторным клеткам органов (экстранейрональный транспорт моноаминов, ЕМТ). Незначительная часть поступившего норадреналина инактивируется под действием двух ферментов: катехол-О-метилтрансфераза (КОМТ; в цитозоле клеток органов) превращает его в норметанефрин, а моноаминоксидаза (МАО; в митохондриях нервных клеток и клеток органов) — в дигидроксиминдальную кислоту.
146 |
147 |

ПРИЛОЖЕНИЕ 29
ПРИЛОЖЕНИЕ 29. МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ВПСП И ТПСП НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ
Последствия взаимодействия медиатора с рецепторами постсинаптической клетки.
А — открытие возбуждающим медиатором Na+-каналов постсинаптического нейрона с его деполяризацией и генерацией в нём потенциала действия;
Б — открытие ингибируюшим медиатором Сl– -каналов постсинаптического нейрона с его гиперполяризацией, а — везикулы ГАМК или глицина, б — рецепторы.
Возбуждающий постсинаптический потенциал (ВПСП). Взаимо-
действуя с рецептором, возбуждающий медиатор (глутамат) открывает неспецифические ионные каналы на постсинаптической мембране клетки так, что повышается их способность к проводимости для одновалентных катионов. Существует сильная движущая сила для натрия, и ионы Na+
устремляются внутрь клетки и деполяризуют ее мембрану. (Следует отметить, что канал проходим и для ионов K+, для которых сохраняется незначительный электрохимический градиент, направленный из внутриклеточной области к внеклеточной среде. Таким образом, через постсинаптическую мембрану одновременно проходят и ионы K+, тем самым слегка противодействуя деполяризации, обусловленной входящим током ионов Na+). Работа каналов ведет к базовому входящему току положительных ионов и, следовательно, к деполяризации постсинаптической мембраны, которая и называется ВПСП. На концевой пластинке нервно-мышечного синапса ВПСП называют также потенциалом концевой пластинки.
Вмеханизме генерации ВПСП участвуют и другие ионные каналы с другими свойствами (лиганд-управляемые), и проводимость этих каналов зависит только от количества молекул трансмиттера, связавшихся с молекулами рецептора. Амплитуда ВПСП лежит в диапазоне от 100 μВ до (в некоторых случаях) 10 мВ. В зависимости от вида синапса, общая продолжительность ВПСП лежит в диапазоне от 5 до 100 мс.
Взоне синапса образовавшийся локально ВПСП распространяется пассивно электротонически по всей постсинаптической мембране клетки. Этот процесс не подчиняется закону «все или ничего». Если большое число возбуждающих синапсов активируются одновременно, то возникает явление суммации, которое реализуется в виде возникновения ВПСП существенно большей амплитуды, что может деполяризовать мембрану всей постсинаптической клетки. Если величина этой деполяризации достигает в области постсинаптической мембраны порогового значения (10 мВ или больше), то на аксонном холмике нервной клетки молниеносно
ПРИЛОЖЕНИЕ279
открываются потенциал-управляемые Na+-каналы, и клетка генерирует потенциал действия, который проводится вдоль аксона. В случае концевой пластинки это приводит к мышечному сокращению. От начала ВПСП до образования потенциала действия проходит еще около 0,3 мс, так что при обильном освобождении транс- миттерапостсинаптическийпотенциалможетпоявитьсяужечерез0,5-0,6мспосле пришедшего в пресинаптическую область потенциала действия. «Синаптическая задержка», выраженная во времени между возникновением пре- и постсинаптического потенциала действия, всегда зависит от типа синапса.
А — процессы освобождения трансмиттера и формирование ВПСП после электрического возбуждения (стрелка), поступившего на пресинаптическое окончание, связаны с возникновением возбуждающего постсинаптического тока (ВПСТ). Если ВПСП переходит порог, то возникает потенциал действия, который благодаря потенциал-управляемым Na+каналам деполяризует мембрану. Б - процессы освобождения трансмиттера и формирование ТПСП связаны с ингибиторным постсинаптическим током (ТПСТ). При этом ток вызывает движение через мембрану ионов К+ (ионы Cl- могут играть роль, если потенциал мембраны позитивнее, чем потенциал равновесия для Cl-, который лежит от -75 до -70 мВ). В — токи мембраны при активации возбуждающих (ВПСТ) и ингибиторных синапсов (ТПСТ) и возникающие в этом случае результирующие постсинаптические потенциалы (ВПСП, ТПСП). При одновременной активации возбуждающих и тормозных синапсов возникающие мембранные токи суммируются, поэтому результирующий постсинаптический потенциал становится маленьким.
148 |
149 |
ПРИЛОЖЕНИЕ 29
!
Тормозной постсинаптический потенциал (ТПСП). На одних тормозных си-
напсах находятся каналы для K+, на других есть и каналы для Cl-. При связывании тормозного трасмиттера (ГАМК, глицин) с рецептором в метаботропном синапсе на постсинаптической мембране повышается проводимость для ионов К+. При нормальной величине потенциала мембраны это приводит к дальнейшему выходящему току ионов К+ и гиперполяризации потенциала мембраны. Возникает так называемыйТПСП.Гиперполяризующиймембранутоктакжесвязансионамихлора: Cl- течет в клетку и гиперполяризует ее.
ПРИЛОЖЕНИЕ 30
ПРИЛОЖЕНИЕ 30. МОТОРНЫЕ БЕЛКИ, ОБЕСПЕЧИВАЮЩИЕ ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ.
АКСОНАЛЬНЫЙ ТРАСПОРТ
В клетках эукариот транспорт и локализация органелл, мембранных везикул, надмолекулярных комплексов, а также отдельных молекул осуществляется вдоль микротрубочек и актиновых микрофиламентов, образующих в них цитоскелет. Транспорт вдоль структур цитоскелета происходит при помощи специализированных механохимических АТФаз, называемых моторными белками. Моторные белки, связанные с микротрубочками, относятся к двум большим семействам – кинезинам и динеинам (белки, связанные с актиновыми микрофиламентами, относятся к миозинам).
Микротрубочкицитоскелета–этонитчатыеневетвящиесяструктурытолщиной 25 нм, состоящие из белков-тубулинов и ассоциированных с ними белков. Микротрубочки являются очень динамичными структурами, которые могут достаточно быстро возникать и разбираться. Роль цитоплазматических микротрубочек может быть сведена к двум функциям: скелетной и двигательной. Скелетная (каркасная) роль заключается в стабилизации формы клетки. Двигательная функция микротрубочек заключается как в создании векторной системы движения, так и в том, что в ассоциации со специфическими ассоциированными моторными белками микротрубочки образуют АТФазные комплексы, способные приводить в движение клеточные компоненты.
Важно, что в цитоскелете микротрубочки образуют радиально расходящиеся поляризованные фибриллы, плюс-концы которых направлены от центра клетки к периферии. Соответственно различают плюс- и минус-направленные моторные белки (кинезины и динеины): они создают возможность для переноса в клетке её компонентов как от периферии к центру (эндоцитозные вакуоли, рециклизация вакуолей ЭПР и аппарата Гольджи), так и от центра к периферии (вакуоли ЭПР, лизосомы, секреторные вакуоли).
Аксоны нейронов могут иметь большую длину (как у гигантских аксонов кальмара) и заполнены большим числом микротрубочек и нейрофиламентов. В аксонах возможно перемещение компонентовклеткикаквнаправлении от сомы к нервному окончанию (антероградный транспорт), так и в противоположном направлении
(ретроградный транспорт).
Аксональный транспорт осуществляется с относительно высокой скоростью: до 400 мм в сутки.
150 |
151 |

ПРИЛОЖЕНИЕ3013
Моторный белок кинезин имеет молекулярную массу около 300 тыс. Да, состоит из двух сходных тяжелых полипептидных цепей и нескольких легких. Каждая тяжелая цепь образует глобулярную головку, которая при ассоциации с микротрубочкойобладаетАТФазнойактивностью,втовремякаклегкиецеписвязываютсяс мембраной пузырьков или других частиц. При гидролизе АТФ изменяется конформация молекулы кинезина и генерируется перемещение частицы в направлении к плюс-концу микротрубочки (антероградный транспорт). Существует целое семейство кинезинов, обладающих сходными моторными головками, но отличающихся хвостовыми доменами. Кинезины связываются с мембранами через мембранные белковые комплексы - кинектины.
За ретроградный транспорт в аксоне отвечает другой белок - цитоплазматический динеин. Он состоит из двух тяжелых цепей - головок, взаимодействующих с микротрубочками, нескольких промежуточных и легких цепей, которые связываются с мембранными вакуолями. Цитоплазматический динеин является моторным белком, переносящим грузы к минус-концу микротрубочек. Динеины делятся на два класса: цитозольные, участвующие в переносе вакуолей и хромосом, и аксонемные, отвечающие за движение ресничек и жгутиков.
Таким образом, в транспорте определенных органелл в клетке могут участвовать сразу несколько различных моторных белков, и в настоящий момент остается открытым научный вопрос координации их действия.
ПРИЛОЖЕНИЕ 31
ПРИЛОЖЕНИЕ 31. ОКСИД АЗОТА (NO) КАК ПАРАКРИННЫЙ МЕДИАТОР
Оксид азота (NO) - это |
|||
паракринный медиатор, вы- |
|||
деляемый |
эндотелиальными |
||
клетками и некоторыми ней- |
|||
ронами. NO быстро окисля- |
|||
ется, поэтому его биологи- |
|||
ческая жизнь |
длится |
всего |
|
несколько секунд (это и есть |
|||
причина его исключительно |
|||
паракринного эффекта). NO |
|||
стимулирует |
растворимую |
||
гуанилатциклазу в клетках- |
|||
мишенях и тем самым по- |
|||
вышает |
внутриклеточную |
||
концентрацию |
цГМФ, |
что, |
|
в свою очередь, стимулиру- |
|||
ет цГМФ-зависимую проте- |
|||
инкиназу. Оксид азота легко |
|||
распространяетсяпутемдиф- |
|||
фузии и может принимать на |
|||
себя функции трансмиттера. |
|||
При этом, например, преси- |
|||
наптические процессы могут |
|||
регулироваться с помощью обратной связи. Образование NO катализируется NO- |
|||
синтазой (NO ) - Са2+-кальмодулин-зависимым ферментом, ускоряющим превра- |
|||
S
щение аргинина в цитруллин и NO. Повышение цитозольного уровня Ca2+ приво-
дит к увеличению образования и выделения NO.
Некоторые физиологические функции оксида азота. NO участвует в процес-
сах долговременной синаптической потенциации, связанной с образованием памяти. В мыслительной деятельности окись азота является и непосредственным участником, и косвенным регулятором. Известно, что NO освобождается нервными окончаниями гранулярных клеток мозжечка и действует на постсинаптические клетки Пуркинье мозжечка. NO выделяется эндотелиальными клетками под влиянием агонистов типа ацетилхолина, присоединение которого к мускариновым рецепторам повышает внутриклеточный уровень Са2+. Освобождение NO эндотелиальнымиклеткамивызываетвазодилатациювблизлежащихклеткахгладкихмышц сосудов. Действуя на гладкомышечные клетки желудочно-кишечного тракта, NO подавляетихсократительнуюактивностьитакимобразомучаствуетвобеспечении моторной функции, при этом некоторые заболевания ЖКТ (болезнь Гиршпрунга, ахалазия кардии) обусловлены снижением числа внутрикишечных нейронов, вырабатывающих NO. Оксид азота активно участвует в почечных физиологических процессах: регулирует почечную гемодинамику, гломерулярную фильтрацию, ингибирует транспорт Na+ и увеличивает его экскрецию.
152 |
153 |
ПРИЛОЖЕНИЕ 13 |
|
ПРИЛОЖЕНИЕ 31 |
154 |
155 |

ПРИЛОЖЕНИЕ 13
В.Ф. Пятин, Г.М. Баишева, И.В. Широлапов, Е.Н. Веретельник
Молекулярная физиология
Учебное пособие для преподавателей и студентов медицинских вузов и биологических факультетов.
Издание 2-е, переработанное, дополненное.
Подписано в печать 18.10.2016. Заказ 1. Формат 60х90/16. Усл. печ. листов 9,75 Издательство ООО «АВИД»
п/а 443063, г. Самара, ул. Ставропольская, 45. Тираж 500 экз.
УДК 576.32/.36
ISBN 978-5-9909123-0-4
Самара, 2016
156