

Історія розвитку цитології. гістології та ембріології.
Будова світлового мікроскопа: механічна, оптична, освітлювальна системи.
Спеціальні метоли світлооптичної мікроскопії: метод "темного поля", поляризаційна мікроскопія, фазово-контрастна мікроскопія.
Спеціальні методи світлооптичної мікроскопії: інтерференційна мікроскопія. люмінісиентна мікроскопія, ультрафіолетова мікроскопія. конфокальна світлова мікроскопія.
Електронна мікроскопія та її види.
Основи гістологічної техніки
гістологічна техніка— сукупність методів і прийомів для вивчення за допомогою мікроскопа хім. складу, фіз. властивостей, життєдіяльності і будови біологічних об'єктів. При вивченні живих об'єктів для підвищення контрастності зображення в мікроскопі використовують фазово-контрастний пристрій, конденсор темного поля (див. Ультрамікроскоп), поляризаційну, інтерференційну мікроскопії та ультрафіолетову мікроскопію. Досліджуючи зміни будови клітин під час їхнього поділу, рухи тощо, використовують мікрокінознімання. При вивченні живих і неживих об'єктів застосовують люмінесцентну мікроскопію з використанням флуоресцентних барвників. Для одержання т. з. постійних (неживих) препаратів об'єкти фіксують (див. Фіксація у біології), заморожують або обезводнюють та заливають у парафін, целоїдин, епоксидну смолу; з непрозорих об'єктів на мікротомі (для електронної мікроскопії — на ультрамікротомі) виготовляють тонкі зрізи. Щоб підвищити контрастність постійних препаратів, застосовують барвники (особливо широко — в гістохімії), які вибірково забарвлюють клітинні структури. Для збільшення контрастності об'єктів, що досліджуються під електронним мікроскопом, їх здебільшого вкривають тонким шаром металевого пилу, застосовують "електронні 6арвники> (чотириокис осмію, ураніл тощо). З метою досягнення негативного контрасту об'єкт вміщують в розчин щільнішої речовини (напр., фосфорно-вольфрамової кислоти), яка заповнює проміжки між частинками, що вивчаються. В. О. Майський
Методика виготовлення постійних та тимчасових гістологічних препаратів.
Постійні цитологічні препарати виготовляють за певною схемою, яка передбачає фіксацію матеріалу та його зневоднення для попередження розкладання вихідної тканини. Для виготовлення постійних цитологічних препаратів застосовують спеціальну методику, яка включає ряд етапів.
Після взяття матеріалу (шматочок тканини завтовшки 1-10 мм) його фіксують, щоб запобігти розкладанню. Для цього використовують 5-20 %-вий розчин формаліну або різні спирти чи складні суміші. Для виготовлення тонких зрізів (завтовшки 5-30 мкм) матеріал спочатку ущільнюють шляхом заморожування або просочування речовинами, які, ущільнюючись самі, роблять тканину придатною для різання.
Ріжуть на спеціальному апараті — мікротомі, який дає можливість отримати тонкі зізи. Потім зрізи фарбують і поміщають між двома скельцями в спеціальне середовище, в якому вони зберігаються.
Слід зазначити, що через слабку контрастність структур нативні гістологічні препарати складно досліджувати, тому для їх успішного вивчення необхідно підвищити контрастність структур. Для цього використовується методика фарбування об’єктів гістологічними барвниками, які елективно (вибірково) забарвлюють лише певні утвори, наприклад ядра, еластичні мембрани, глікоген, ліпіди. Залежно від того, які фарби сприймає структура, останню називають базофільною (якщо вона фарбується основними барвниками) чи оксифільною (при -забарвленні кислими фарбами). Метахрізмазія — це явище, коли певний утвір фарбується не в колір фарби, а в якийсь інший, тобто, коли між хімічними речовинами клітини і фарбою утворюються сполуки, що мають особливе забарвлення, не схоже на застосовану фарбу.
8 Основні методи дослідження, які застосовуються в гістології, ембріології та цитології.
Гістологічні методи дослідження застосовуються для вивчення будови і функції клітин і тканин в нормі, патології та експерименті. Основою гістологічного методу дослідження є гістологічна техніка - комплекс методичних прийомів, використовуваних при виготовленні препаратів клітин і тканин для їх мікроскопічного дослідження. Мікроскопічне вивчення клітин і тканин може проводитися двома основними шляхами в залежності від стану досліджуваного об'єкта: дослідження живих клітин і тканин, дослідження неживих клітин і тканин, що зберігають структуру завдяки спеціальним прийомам фіксації.
Вивчення живих об'єктів дає можливість спостерігати фізіологічні процеси в клітинах ітканинах, їх прижиттєве будову. Воно проводиться на клітинах, вільно зважених в рідкому середовищі (клітинах крові, епітеліальних клітинах зіскрібків і ін), а також на культурах клітин і тканин, вирощених на спеціальних поживних середовищах. Об'єктом прижиттєвого спостереження можуть бути тонкі, прозорі тканинні плівки (брижа, плавальна перетинка).Широке застосування живих об'єктів обмежена великими технічними труднощами, пов'язаними з властивостями переживають тканин. Частіше використовується фіксований матеріал.
Мета фіксації зберегти прижиттєву структуру клітин і тканин шляхом швидкого впливу на них хімічними агентами, що запобігають розвиток посмертних змін. Вибір методу фіксації залежить від завдань дослідження і особливостей фіксованої матеріалу.
При гістологічному дослідженні, спочатку роблять зріз тканини, потім шматочок тканини зневоднюють спиртом, після чого заливають у ущільнюючі середовища - парафін, целлоидин.Заливка парафіном дозволяє отримати більш тонкі зрізи. Приготовлені зрізи забарвлюють для чіткого виділення структур клітин і тканин, які по-різному сприймають барвники.
Приготування гістологічного препарату завершується укладенням його в середовища, що забезпечують збереження структур об'єкта, його забарвлення і прозорості. Найбільш часто для цих цілей застосовують органічні смоли.
Цитохімічні дослідження засновані на використанні специфічних хімічних кольорових реакцій для визначення в клітинах різних речовин (під дією спеціально підібраних реактивів відбувається фарбування тих чи інших речовин у цитоплазмі, а за ступенем і характером забарвлення судять про кількість або активності досліджуваних речовин). Цитохімічні дослідження щодо нескладні, але поступаються у точності кількісному аналізу, що проводиться з допомогою біохімічних методів.
При цитохімічним дослідженні частіше користуються напівкількісної оцінкою результатів, використовуючи принцип Астальді, заснований на виявленні різного ступеня інтенсивності специфічного забарвлення. У залежності від неї досліджувані елементи ділять на 4 групи: з негативною реакцією (-), слабоположительная (+), позитивної (+ +) і різко позитивної (+++).Для кількісного вираження результатів підраховують 100 клітин певного виду, диференціюючи їх за вказаним принципом, потім число клітин з однаковою інтенсивністю забарвлення множать на відповідне цій групі число плюсів, сума цих творів становить умовні одиниці. Наприклад, при дослідженні активності лужної фосфатази в нейтрофілах з 100 переглянуто клітин в 60 клітинах активність ферменту не виявлено (-), у 35 - специфічна забарвлення була слабкою (+) і в 5 - більш інтенсивної (++). Результат визначення активності лужної фосфатази в нейтрофілах у такому випадку складе (60 * 0) + (35 * 1) + (5 * 2) = 0 +35 +10 = 45 од.
Можна висловити результат у вигляді середнього цитохімічного показника по L. Kaplow (1955) або середнього цитохімічного коефіцієнта (СЦК). З цією метою також диференціюють 100 досліджуваних клітин за вказаною вище системі. Отриманий відсоток клітин в кожній групі множать на відповідне цій групі число плюсів. Сума цих величин, поділена на 100, являє собою СЦК для однієї клітини. У зазначеному прикладі СЦК лужної фосфатази нейтрофілів дорівнює 0,45.
У тих випадках, коли досліджувані речовини локалізуються в клітинах у вигляді одиничних гранул (наприклад, активність неспецифічної естерази у лімфоцитах і ін), результат цитохімічних реакцій доцільно виражати у відсотках клітин, що дають позитивну реакцію.
Метод напівкількісної оцінки є орієнтовними, але дозволяє порівнювати розподіл досліджуваних речовин у різних клітинних елементах або в одних і тих же клітинах при різнихпатологічних станах організму, а також залежно від перебігу захворювання, ступеня його важкості та у зв'язку із проведеною терапією.
Слід мати на увазі, що цитохимический метод може бути використаний тільки як доповнення до морфологічному дослідженню, але не може його замінити. Недоліком всіх цитохімічних реакцій є їх приблизна якісна оцінка, заснована на ступені інтенсивності забарвлення.
Загальні уявлення про клітину та її структуру.
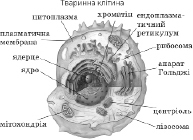
Клітина (cellula, cytus) – основна структурно-функціональна одиниця всіх живих організмів, елементарна жива система. Може існувати як окремий організм (бактерії, простіші, деякі водорості, гриби) або в складі тканин багатоклітинних тварин, рослин, грибів. Лише віруси є неклітинною формою життя.( Рис.1)
Вміст клітини – протоплазма. В кожній клітині є генетичний апарат, який в клітинах еукаріот міститься в ядрі, відокремлений мембранами від цитоплазми, а в клітинах прокаріотів, які не мають оформленого ядра – в нуклеоїді.
- Клітини еукаріотів здатні до самоутворення шляхом мітозу.
- Статеві клітини утворюються шляхом мейозу.
- Розміри клітини коливаються від 0,1-0,25
мкм (деякі бактерії) до 155 мм (яйце страуса
в шкарлупі) .
Розміри клітини коливаються від 0,1-0,25
мкм (деякі бактерії) до 155 мм (яйце страуса
в шкарлупі) .
- Діаметр більшості еукаріотних клітин лежить у межах 10-100 мкм.
- Різноманітні функції в клітині виконуються за допомогою органоїдів.
- Найважливіші хімічні компоненти клітини: білки, включаючи ферменти, містяться як у клітинах, так і в рідких середовищах, але синтезуються лише в клітині.
Рис. 1. Форма клітин людини і тварин: а - нервова клітина (нейрон); б, в, г — різні види епітеліальних тканин, д-клітина сполучної тканини; е — яйцеклітина; є — м'язова клітина.
- Для клітин є характерним принцип компактності (особливо виражений у структурі ДНК).
- Всередині клітини безперервно підтримується концентрація іонів, яка відрізняється від концентрації іонів у міжклітинному просторі.
- Клітинам властивий фагоцитоз (втягування клітинної мембрани і замикаються, захоплення клітинами великих молекул, білків, вірусів).
- Клітини рослин поверх клітинної мембрани покриті твердою клітинною оболонкою, яка має пори; коли клітини припиняють свій ріст, оболонки часто просочуються лігнином, кремнеземом, що надає стійкості рослині (механічні тканини).
- Клітини рослин, які високо диференційовані, мають 1 чи 2 центральні вакуолі (запас солей і води) і хлоропласти.
- Всі клітини еукаріотів мають схожий набір органоїдів.
- Різні клітини одного організму відрізняються формою і розміром, хімічним складом, функціями.
- Клітини в багатоклітинному організмі мають різну спеціалізацію ( нервові, м‘язові, залозеві (в них синтезуються гормони і ферменти).
- Багато клітин поліфункціональні (клітини печінки синтезують різні білки і жовч, накопичують глікоген і перетворюють його у глюкозу, окислюють невластиві їй речовини).
- Клітини близької за походженням і функцією, які створюють тканини (гістогенез).
- В клітинах постійно підтримуються процеси саморегуляції (обмін речовин або метаболізм).
- В організмі людини ≈ 1014 клітин. В деяких тканинах кількість клітин постійна протягом життя.
- у людини кожного дня гине ≈ 70 млрд клітин кишкового епітелію і 2 млрд еритроцитів.
- Мінімальна тривалість життя клітини людини 1-2 дні (кишковий епітелій).
- Клітини відрізняються за формою:
1) циліндричні і кубічні (в епітелії);
2) дископодібні (еритроцити), кулясті (яйцеклітини);
3) видовжені і веретеноподібні (м‘язові);
4) зірчасті (нервові).
Клітинна теорія. Історія створення.
Клітинну теорію в 1838–1839 роках сформулювали ботанік Матіас Шлейден і зоолог Теодор Шванн. Ці вчені довели принципову подібність між собою тваринних та рослинних клітин, і на основі всіх накопичених до того часу знань постулювали, що клітина є структурною та функціональною одиницею всіх живих організмів. 1855 року Рудольф Вірхов доповнив клітинну теорію твердженням лат. «Omnis cellula ex cellula» — «Кожна клітина — з клітини».
Клітинна теорія є однією із основоположних ідей сучасної біології, вона стала незаперечним доказом єдності всього живого та фундаментом для розвитку таких дисциплін як ембріологія, гістологія та фізіологія. Основні положення клітинної теорії не втратили своєї актуальності, проте від часу створення її було доповнено, і наразі вона містить такі твердження:
Клітина — елементарна одиниця будови, функціонування, розмноження і розвитку всіх живих організмів, поза межами клітини немає життя.
Клітина — цілісна система, що містить велику кількість пов'язаних один з одним елементів — органел.
Клітини різних організмів схожі (гомологічні) за будовою та основними властивостями і мають спільне походження.
Збільшення кількості клітин відбувається шляхом їх поділу, після реплікації її ДНК: клітина — від клітини.
Багатоклітинний організм — це нова система, складний ансамбль із великої кількості клітин, об'єднаних та інтегрованих у системи тканин і органів, пов'язаних між собою за допомогою хімічних факторів: гуморальних і нервових.
Клітини багатоклітинних організмів мають однаковий набір генетичної інформації, але відрізняються за рівнем експресії(роботи) окремих генів, що призводить до їх морфологічної та функціональної різноманітності — диференціації.
Слід зазначити, що в різних джерелах кількість та формулювання окремих положень сучасної клітинної теорії можуть відрізнятись.
Більшість еукаріотичних клітин мають розміри до 100 мкм, а прокаріотичні ще на порядок менші, тому людина не може бачити їх неозброєним оком. Відкриття та дослідження клітин стало можливим тільки після винайдення Янсеном оптичного мікроскопа (1590 року).
1665 року, вивчаючи будову корка під мікроскопом, Роберт Гук вперше помітив, що тканина живого організму, складається із маленьких комірок. Ці комірки він назвав «клітинами». Гук припускав, що клітини порожні, а живою речовиною є клітинні стінки[1]. Його дослідження стали поштовхом для систематичного вивчення анатомії рослин, зокрема такими вченими як Мальпігіта Грю. Їхні результати підтвердили висновки Гука про те, що тіло рослин складається із щільно розміщених комірок.
Мікроскоп, який використовував Роберт Гук, давав збільшення тільки до 30X, що робило майже не можливим вивчення внутрішньої будови клітин. У другій половині XVII століття торговцю тканинами Антоні ван Левенгуку вдалось змайструвати кращий однолінзовий мікроскоп із збільшенням 300X. З його допомогою Левенгук спостерігав живі клітини, зокрема одноклітинніводорості і найпростіших із ставкової води, бактерії, людські еритроцити та сперматозоїди. Свої відкриття він описав у ряді повідомлень до Лондонського королівського товариства.
Подальше дослідження клітин обмежувалось двома факторами: по-перше мікроскопи у XVIII столітті мали порівняно невелику роздільну здатність, по-друге біологія в той час мала переважно описовий, а не експериментальний характер. Тому нові досягнення в цій галузі були зроблені аж у 30-их роках XIX століття, коли почали використовуватись дволінзові мікроскопи. Використовуючи такий прилад англійський ботанік Роберт Браун відкрив 1833 року ядро, як сферичне тільце, наявне в рослинних клітинах. Ян Пуркіньє встановив, що живим компонентом клітини є внутрішній вміст, який він назвав «протоплазмою».
У 1838 році ботанік Матіас Шлейден дійшов важливого висновку, що всі рослинні тканини складаються із клітин, а зародки рослин завжди розвиваються із однієї клітини. Роком пізніше німецький цитолог Теодор Шванн поширив аналогічні висновки і на тканини тварин. Таким чином він став першим, хто встановив фундаментальну схожість між рослинними та тваринними тканинами. На основі накопичених спостережень Шванн створив клітинну теорію, згідно з якою клітина є основною структурною та функціональною одиницею живих організмів.
Через 20 років клітинна теорія була доповнена ще одним важливим принципом, встановити який у великій мірі вдалось завдяки дослідженням клітинного поділу Карлом Негелі. 1855 рокуРудольф Вірхов довів, що всі клітини утворюються із інших клітин шляхом поділу. Таким чином, клітина була встановлена роль клітини як одиниці розмноження живих організмів. До кінця XIX століття було описано всі структури клітини, які можна було вивчати за допомогою оптичного мікроскопа. І тільки у 1950-их роках, коли Паладе, Протер та Шестранд, розробили методи фіксації і фарбування біологічних зразків для електронної мікроскопії, стало можливим вивчення ультрастукртури клітини
У формуванні сучасної клітинної біології, крім цитології, що зосереджується в першу чергу на будові клітини та її компонентів, важливу роль відіграли такі галузі біологічної науки як біохімія та генетика. Внаслідок стрімкого розвитку цих дисциплін у XX столітті уявлення про життєдіяльність клітин були значно розширені.
Хімічні елементи у клітині.
М
 ал.
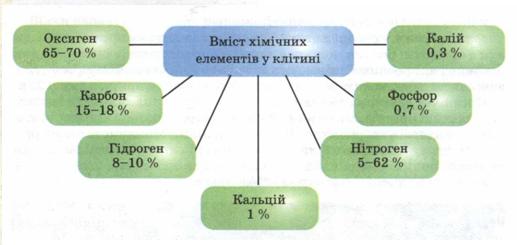
Вміст хімічних елементів у клітині
ал.
Вміст хімічних елементів у клітині
У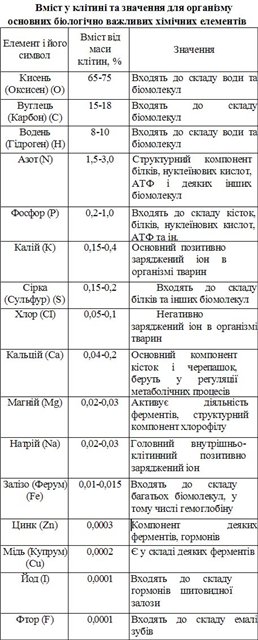 складі живих істот виявлено понад 60
хімічних елементів. Хімічні елементи,
що постійно входять до складу організмів
і необхідні для їхньої життєдіяльності,
називають біогенними. Понад 90 % вмісту
клітин становлять такі важливі біогенні
елементи, як Оксиген, Карбон, Гідроген,
Нітроген. Серед інших важливе значення
мають Кальцій, Калій, Фосфор, Магній,
Сульфур, Ферум, Купрум, Хлор, Натрій
(мал. 20). Ці біогенні елементи універсальні.
Вони є у клітинах усіх видів організмів.
Вміст інших хімічних елементів значно
нижчий, вони можуть траплятися в
організмів одних видів і не траплятися
в інших.
складі живих істот виявлено понад 60
хімічних елементів. Хімічні елементи,
що постійно входять до складу організмів
і необхідні для їхньої життєдіяльності,
називають біогенними. Понад 90 % вмісту
клітин становлять такі важливі біогенні
елементи, як Оксиген, Карбон, Гідроген,
Нітроген. Серед інших важливе значення
мають Кальцій, Калій, Фосфор, Магній,
Сульфур, Ферум, Купрум, Хлор, Натрій
(мал. 20). Ці біогенні елементи універсальні.
Вони є у клітинах усіх видів організмів.
Вміст інших хімічних елементів значно
нижчий, вони можуть траплятися в
організмів одних видів і не траплятися
в інших.
Усі біогенні елементи, незалежно від їхнього вмісту, впливають на життєдіяльність організмів. За відсутності того чи іншого хімічного елемента можуть порушуватись процеси життєдіяльності або істота взагалі гине. Наприклад, за відсутності Магнію та Феруму листки рослини стають блідо-зеленими або жовтіють і процес фотосинтезу гальмується. Це пояснюється тим, що без цих хімічних елементів не може утворюватись пігмент хлорофіл.
Молекулярний склад клітини.
На молекулярному рівні хімічні сполуки класифікують на:
Неорганічні сполуки — 60-70 % маси тіла;
Вода
Розчинення гідрофобних та гідрофільних речовин;
Терморегуляція;
Транспорт речовин;
Гідроліз та окиснення високомолекулярних речовин;
Підтримання об'єму, тургору та пружності клітини;
Мінеральні солі:
Субстрат катіонів та аніонів;
Регуляторна функція;
Органічні сполуки — 15-25 % маси тіла.
Гістохімічні методи виявлення основних груп органічних речовин.
Поверхневі структури клітини.
У складі поверхневого апарату клітини виділяють 3 компоненти:
1. Плазматичну мембрану, або плазмолеммой
2. Надмембранний комплекс, або глікокаликс
3. Субмембранний комплекс або субмембранний опорно-скорочувальний апарат.
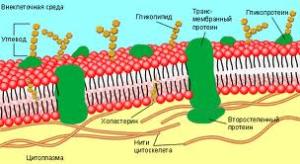
Плазмолемма - є структурною і функціональною основою поверхневого апарату клітини і являє собою сферично замкнуту біомембранами. Структура плазмолеми відповідає рідинно-мозаїчної моделі мембран.
Надмембранний комплекс, або глікокаликс є наружней частиною поверхневого апарату клітини, розташовуючись над плазмолеммой.
Поверхневий апарат клітин складається зі структур, розташованих над плазматичною мембраною (над-мембранні комплекси), самої мембрани, а також деяких структур, розташованих під нею (підмембранні комплекси). Що таке надмембранні комплекси клітин? У клітинах прокаріотів, грибів і рослин плазматична мембрана ззовні вкрита клітинною стінкою. У рослин вона складається переважно з нерозчинних у воді, зібраних у пучки волокон целюлози. Ці волокна утворюють каркас, заглиблений в основу (матрикс). Матрикс також складається здебільшого із полісахаридів. Залежно від типу тканин рослин і функцій, які вони виконують, до складу клітинної стінки також можуть входити й інші сполуки: ліпіди, білки, неорганічні сполуки (двооксид силіцію, солі кальцію тощо).
Ч ерез
клітинні стінки рослин відбувається
транспорт води і певних сполук. Проникність
стінок рослинних клітин можна
проілюструвати на прикладі явищ
плазмолізу і деплазмолізу. Наприклад,
якщо клітину вмістити у розчин з
концентрацією солей, вищою, ніж у
цитоплазмі,
то вода виходитиме з неї. Це спричиняє
явище плазмолізу (від грец. плазма
-виліплення, утворення та лізіс —
розчинення) - відшарування пристінкового
шару цитоплазми від клітинної стінки.
Якщо ж клітину вмістити у розчин солей
з концентрацією нижчою, ніж у її
цитоплазмі, то спостерігається зворотний
процес: вода надходитиме в клітину,
внаслідок чого зростає внутрішньоклітинний
тиск. Це явище називають деплазмолізом
(від лат. де - префікс, що означає відміну).
У різних груп грибів структура і хімічний
склад клітинної стінки можуть розрізнятись.
Як і у рослин, її основу складають
різноманітні полісахариди (целюлоза,
хітин тощо). До складу клітинних стінок
деяких грибів можуть входити темні
пігменти (меланіни), амінокислоти,
фосфати та інші сполуки.
ерез
клітинні стінки рослин відбувається
транспорт води і певних сполук. Проникність
стінок рослинних клітин можна
проілюструвати на прикладі явищ
плазмолізу і деплазмолізу. Наприклад,
якщо клітину вмістити у розчин з
концентрацією солей, вищою, ніж у
цитоплазмі,
то вода виходитиме з неї. Це спричиняє
явище плазмолізу (від грец. плазма
-виліплення, утворення та лізіс —
розчинення) - відшарування пристінкового
шару цитоплазми від клітинної стінки.
Якщо ж клітину вмістити у розчин солей
з концентрацією нижчою, ніж у її
цитоплазмі, то спостерігається зворотний
процес: вода надходитиме в клітину,
внаслідок чого зростає внутрішньоклітинний
тиск. Це явище називають деплазмолізом
(від лат. де - префікс, що означає відміну).
У різних груп грибів структура і хімічний
склад клітинної стінки можуть розрізнятись.
Як і у рослин, її основу складають
різноманітні полісахариди (целюлоза,
хітин тощо). До складу клітинних стінок
деяких грибів можуть входити темні
пігменти (меланіни), амінокислоти,
фосфати та інші сполуки.
У прокаріотів структура клітинної стінки досить складна. У більшості бактерій вона складається з високомолекулярної сполуки - муреїну, який надає їй міцності. До складу клітинної стінки бактерій також входять білки, сполуки ліпідів з полісахаридами тощо (мал. 45).
Мал. 45. Схема будови клітинної оболонки бактерії: 1 - плазматична мембрана; 2 - клітинна стінка; З - білки; 4 - муреїн
Залежно від будови і властивостей надмембранних комплексів бактерій поділяють на грампозитивних і грамнегативних. У грамнегативних бактерій структура надмембранних комплексів складніша завдяки шару полісахаридів і додатковій зовнішній мембрані. Тому на них не діють певні антибіотики (наприклад, пеніцилін, актиноміцин). Клітинна стінка бактерій має антигенні властивості, що дає можливість певним групам лейкоцитів «впізнавати» хвороботворні бактерії і виробляти до них антитіла.
Будова та функції плазматичної мембрани.
П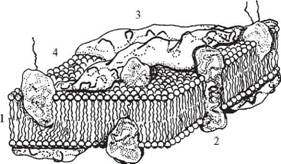 лазматична
мембрана, або плазмалема,
- найбільш постійна, основна, універсальна
для усіх клітин мембрана. Вона є найтоншою
(близько 10 нм) плівкою, що покриває усю
клітину. Плазмалема складається з
молекул білків і фосфоліпідів.
лазматична
мембрана, або плазмалема,
- найбільш постійна, основна, універсальна
для усіх клітин мембрана. Вона є найтоншою
(близько 10 нм) плівкою, що покриває усю
клітину. Плазмалема складається з
молекул білків і фосфоліпідів.
Відповідно до сучасних досліджень мембрана має рідинно-мозаїчну структуру (С. Сінгер і Дж. Нікольсон, 1972) (рис. 2).
Рис- 2- Узагальнена модель типової плазматичної мембрани:
1 — ліпіди; 2 — інтегральні білки; 3 — периферичні білки; 4 — глікопротеїди
В основі мембранної матриці лежить двошарова ліпідна структура. Більшу частку мембранних ліпідів складають фосфоліпіди, які здатні утворювати бішар, що складається з внутрішньої гідрофобної області (аліфатична частина жирних кислот або стероїдного кістяка холестерину) і гідрофільних поверхонь (гліцерин, залишок фосфорної кислоти, аміноспирту, OH-жирної кислоти). Тому через мембрани можуть проникати жиророзчинні речовини, а водорозчинні речовини і гідрофільні іони не в змозі перебороти гідрофобну область; вони попадають усередину клітини по спеціальних каналах проникності білкової природи. Основну частину ліпідів, що містяться в біомембранах, являють собою полярні ліпіди (гліцерофосфоліпіди, сфінгофосфоліпіди, гліцерогліко-ліпіди, сфінгогліколіпіди). Окремі фрагменти молекул фосфоліпідів чинять тепловий рух, що відіграє важливу роль: 1) у взаємодії ліпідів з білками; 2) у процесах транспорту речовин через мембрану; 3) у проникності
Молекули фосфоліпідів розташовані в два ряди - гідрофобними кінцями всередину, гідрофільними голівками до внутрішнього і зовнішнього водного середовища. В окремих місцях біслой (подвійний шар) фосфоліпідів наскрізь пронизаний білковими молекулами (інтегральні білки). Усередині таких білкових молекул є канали - пори, через які проходять водорозчинні речовини.
Плазматична мембрана насамперед захищає внутрішнє середовище клітини від несприятливих впливів і бере участь у процесах обміну речовин із навколишнім середовищем. Вона утворює вирости, мікроворсинки, які значно збільшують поверхню клітини. У плазматичній мембрані розташовані деякі ферменти, необхідні для обміну речовин.
С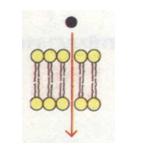 полуки,
потрібні для життєдіяльності клітин,
а також продукти обміну речовин
перетинають плазматичну мембрану за
допомогою дифузії (мал. 3), пасивного чи
активного транспорту. Нагадаймо, що
дифузія (від лат. диффузіо - розлиття) -
процес, за якого речовини проникають
крізь певні ділянки і пори мембран
унаслідок їхньої різної концентрації
по обидва її боки. Цей процес відбувається
без витрат енергії у результаті хаотичного
теплового руху молекул.
полуки,
потрібні для життєдіяльності клітин,
а також продукти обміну речовин
перетинають плазматичну мембрану за
допомогою дифузії (мал. 3), пасивного чи
активного транспорту. Нагадаймо, що
дифузія (від лат. диффузіо - розлиття) -
процес, за якого речовини проникають
крізь певні ділянки і пори мембран
унаслідок їхньої різної концентрації
по обидва її боки. Цей процес відбувається
без витрат енергії у результаті хаотичного
теплового руху молекул.
Мал. 3. Схема транспорту речовин через плазматичну мембрану за допомогою дифузії
В ибіркове
проникнення речовин через мембрани
забезпечує пасивний транспорт (мал. 4).
Для нього, як і для дифузії, характерне
переміщення речовин з боку, де концентрація
вища. Пасивний транспорт забезпечується
за участю рухомих мембранних
білків-переносників; зміною просторової
структури білкій, які перетинають
мембрану; та через канали у мембрані.
ибіркове
проникнення речовин через мембрани
забезпечує пасивний транспорт (мал. 4).
Для нього, як і для дифузії, характерне
переміщення речовин з боку, де концентрація
вища. Пасивний транспорт забезпечується
за участю рухомих мембранних
білків-переносників; зміною просторової
структури білкій, які перетинають
мембрану; та через канали у мембрані.
Активний транспорт речовин через біологічні мембрани пов'язаний із витратами енергії, оскільки не залежить від концентрації речовин, які мають потрапити в клітину або вийти з неї (мал. 4). На цей процес впливає різниця концентрацій іонів калію і натрію у зовнішньому середовищі та всередині клітини. Тому його назвали калій-натріевим насосом. Концентрація іонів калію всередині клітини вища, ніж ззовні, а іонів натрію - навпаки. Завдяки цьому іони натрію пересуваються в клітину, а калію - з неї. Але концентрація цих іонів у живій клітині і поза нею ніколи не вирівнюється, оскільки існує особливий механізм, який іони натрію «відкачує» з клітини, а калій - «закачує» в неї. Цей процес потребує витрат енергії.
М ал.
4. Схема пасивного (1) та активного (2)
транспорту речовин через плазматичну
мембрану
ал.
4. Схема пасивного (1) та активного (2)
транспорту речовин через плазматичну
мембрану
Завдяки механізму калій-натрієвого насосу енергетично сприятливе (тобто таке, що сприяє вирівнюванню концентрації) пересування іонів натрію в клітину, полегшує енергетично несприятливий (в бік вищої концентрації) транспорт низькомолекулярних сполук (глюкози, амінокислот тощо).
Процеси дифузії, пасивного і активного транспорту властиві всім типам біологічних мембрани.
Існує ще один механізм транспорту речовин через мембрани, який називають ендоцитозом. Розрізняють два основні види ендоцитозу: фаго- і піноцитоз.
Фагоцитоз (від грец. фагос - пожирати) - це активне захоплення твердих об'єктів мікроскопічних розмірів (частинок органічних сполук, дрібних клітин та ін.) (мал. 5). До фагоцитозу здатні лише певні типи клітин тварин. Адже на відміну від клітин прокаріотів, рослин і грибів, вони позбавлені щільної клітинної стінки. За допомогою фагоцитозу захоплюють їжу деякі одноклітинні (наприклад, амеби, форамініфери) та спеціалізовані клітини багатоклітинних (наприклад, травні клітини гідри) тварин.
Макрофаги за допомогою фагоцитозу здійснюють захисну функцію. Вони захоплюють і перетравлюють сторонні частки і мікроорганізми. Явище фагоцитозу в 1892 p. відкрив видатний український учений І.І. Мечников.
Надмембранний комплекс.
Надмембранний комплекс — це оболонка вуглеводної природи, розміщена над плазматичною мембраною.
Надмембранний комплекс, або глікокаликс є наружней частиною поверхневого апарату клітини, розташовуючись над плазмолеммой.
До складу надмембранний комплексу включають:
1. Вуглеводні частини гліколіпідів і глікопротеїдів
2. Периферичні мембранні білки, розташовані на наружней частини билипидного шару
3. Інтегральні і полуінтегральние білки, що мають зовнішню зону, виступаючу над біліпідного шаром.
4. Специфічні вуглеводи, не пов'язані хімічно з компонентами мембрани, локалізовані над біліпідного шаром.
5. Субмембранний комплекс або субмембранний опорно-скорочувальний апарат - розташовується під плазмолеммой, з внутрішньої сторони поверхневого апарату клітини. До складу субмембранного опорно-скорочувального апарату виділяють периферичну гіалоплазму і опорно-скоротливу систему.
Уявлення про кортикальний шар цитоплазми
Основними складовимигликокаликса служать комплекси полісахаридів з білками (>гликопротеини) і жирами (>гликолипиди). Зсередини до мембрани примикає кортикальний (кортикальний) шар цитоплазми завтовшки 0,1—0,5мкм, у якому невідомі рибосоми і бульбашки, але у значну кількість перебувають микротрубочки імикрофиламенти, які мають у собісократимі білки.
периферійний (кортикальний) шар цитоплазми. В ньому відсутні включення і він зафарбований в рожевий колір. Кортикальний шар цитоплазми обмежений первинною оболонкою (плазмолемою), поверх якої розташована вторинна оболонка яйцеклітини. Вона утворена декількома шарами фолікулярних клітин та їх похідними. Поблизу первинної оболонки вони формують променевий вінець, який включає прозору зону.
Властивості та функції плазматичної мембрани.
Функції плазматичної мембрани
Мембрана є не лише бар'єром між клітиною і зовнішнім середовищем, але являє собою пристрій, що забезпечує відносну сталість складу внутрішньоклітинного об'єму. Поряд з цим вона містить специфічні рецептори для зовнішніх «сигналів», присутністю яких можуть пояснюватися такі різноманітні відповіді, як орієнтований рух клітини (хемотаксис), стимуляція пов'язаних із мембраною ферментів (циклаз) чи генерації сигналів, що можуть бути хімічними (цАМФ, цГМФ) чи електричними, як у нервовій клітині. Плазматична мембрана також є місцем розташування специфічних для клітини антигенів, що характерні як для даного типу клітин, так і для виду ссавця в цілому. Білки і специфічні ферменти, локалізовані в мембрані, пов'язані з транспортом іонів і метаболітів через мембранний бар'єр. Завдяки інвагінаціям (впинанням) плазматична мембрана може утворювати єдине ціле з довгими внутрішньоклітинними каналами, що дозволяє здійснювати надходження речовин ззовні далеко «всередину» клітини, і бере участь у формуванні простору між внутрішньою і зовнішньою мембранами ядра.
Види трансмембранного транспорту.
Усі клітинні мембрани є рухливими текучими структурами, оскільки молекули ліпідів і білків не пов'язані між собою ковалентними зв'язками і здатні досить швидко переміщатися в площини мембрани. Завдяки цьому мембрани можуть змінювати свою конфігурацію, тобто мають плинність.
Мембрани - структури дуже динамічні. Вони швидко відновлюються після ушкодження, а також розтягуються і стискуються при клітинних рухах.
Мембрани різних типів клітин істотно розрізняються як по хімічному складу, так і за відносним змістом в них білків, глікопротеїнів, ліпідів, а отже, і по характеру наявних в них рецепторів. Кожен тип клітин тому характеризується індивідуальністю, яка визначається в основному глікопротеїнами. Розгалужені ланцюги глікопротеїнів, виступаючі з клітинної мембрани, беруть участь в розпізнанні чинників зовнішнього середовища, а також у взаємному пізнаванні споріднених клітин.
Трансмембранний транспорт речовин здійснюється в трифазних системах, які включають дві фази, розділені тонкою мембраною. Керуючи структурою і властивостями мембрани і складових її компонентів, можна здійснювати Високоселективне поділ і очищення речовин. Процеси мембранного транспорту вкрай важливі не тільки в біологічних системах.
Обмін речовин між клітиною і середовищем визначається транспортною функцією ПАК. У своїй діяльності клітина використовує кілька видів транспорту молекул і речовин через ПАК:
Вільний транспорт, або проста дифузія.
Пасивний транспорт, або полегшена дифузія
Активний транспорт
Транспорт в мембранної упаковці або цитоз.
Вільний транспорт - здійснюється тільки за наявності електричного градієнта по обидві сторони мембрани. Цей градієнт існує тільки при різниці концентрації і \ або зарядів транспортуються молекул.
Величина градієнта визначає напрямок і швидкість вільного транспорту. Такий напрям транспорту називають транспортом по градієнту концентрації. При цьому швидкість вільного транспорту прямолінійна величиною градієнта. Транспорт по градієнту концентрації призводить до зменшення різниці концентрацій і поступового зниження швидкості вільного транспорту.
Біологічна роль вільного транспорту обмежена. Це визначається його недостатньою вибірковістю. Через біліпідний шар можуть проходити будь гідрофобні молекули. Більшість біологічно активних молекул є гідрофільними, тому їх вільний транспорт через біліпідний шар утруднений.
Пасивний транспорт - полегшена дифузія - також здійснюється тільки по градієнту концентрацій і без витрат АТФ. Швидкість пасивного транспорту набагато більше, ніж вільного. При збільшенні різниці концентрацій настає момент, коли швидкість стає постійною. Транспорт здійснюється спеціальними молекулами - переносниками. За їх допомогою через мембрану по градієнту концентрації транспортуються великі гідрофільні молекули (цукру, амінокислоти). У ПАК є пасивні переносники для різних іонів (К +, Na +,Ca 2 +, Cl -, HCO 3 -).
Особливістю пасивних переносників є їх висока специфічність (вибірковість) по відношенню до транспортуються молекулам. Друга особливість - висока швидкість транспорту, яка може становити 10 4молекул в секунду і більше. Клітина може регулювати кількісний і якісний набір переносників у своєму ПАК. Це дозволяє клітині диференціюватися і реагувати на зміни умов.
Механізм дії переносників заснований на їх здатності утворювати канали, специфічні для певних молекул. Наприклад: пасивний переносник глюкози.
Змінювати параметри пасивного транспорту в клітині можна за допомогою лікарських препаратів, антибіотиків. Антибіотики виступають в ролі пасивних переносників. У еукаріотічеккіх клітинах порушення пасивного транспорту можуть викликати деякі токсини і отрути. Білки переносники можуть здійснювати і пасивний транспорт і активний транспорт.
Унипортом (Uniport) - транспорт у градієнтному (одному) напрямку однієї речовини.
Симпорт (Symport) - транспорт у градієнтному напрямку декількох речовин.
Антіпорт (Antiport) - транспорт у декількох напрямках декількох речовин.
2. пасивний транспорт
2.1. проста дифузія
2.2. осмос
2.3. дифузія іонів
2.4. полегшена дифузія
3. активний транспорт
3.1. Первинно-активний транспорт
3.2. Вторинно-активний транспорт
Проникність плазматичної мембрани для води та іонів.
Найважливішою властивістю мембрани є також виборча проникність. Це означає, що молекули та іони проходять через неї з різною швидкістю, і чим більше розмір молекул, тим менше швидкість проходження їх через мембрану. Ця властивість визначає плазматичну мембрану як осмотичний бар'єр. Діти, подивіться на малюнок 6 та 7, щоб зрозуміти, процес проникнення (дифузії).
Максимальну проникаючу здатність має вода і розчинені в ній гази; значно повільніше проходять крізь мембрану іони. Дифузія води через мембрану називається осмосом. Давайте роздивимося цей процес на відео.
Природа поляризації клітинних мембран на сьогоднішній день визначена. Вона базується на особливостях будови і функціонування клітинних мембран, що мають виборчу проникність і здатні змінювати проникність у залежності від функціонального стану. Крім того, необхідно враховувати іонний склад позаклітинного середовища і внутрішньоклітинної рідини. Мембрана легко проникна для жиророзчинних речовин, молекули яких проникають крізь бімолекулярний шар ліпідів. Великі водорозчинні молекули — аніони органічних кислот, зовсім не проходять через мембрану, а можуть залишати клітину лише шляхом екзоцитозу. У мембрані також існують канали, проникні для води, малих молекул водорозчинних речовин і малих іонів.є Причина виникнення хвилі збудження - зміна іонної проникності мембрани. При дії подразника проникність клітинної мембрани дляNа+ підвищується, іони натрію дифундують в клітину. Відповідно із зменшенням електропозитивного заряду зовнішньої сторони мембрани зменшується електронегативний заряд внутрішньої сторони мембрани. Відбувається деполяризація мембрани - зменшення МП. У перший момент деполяризація йде повільно, МП зменшується лише на 15-25 мВ. Початкова деполяризація отримала назву - локальна (місцева) відповідь. Деполяризація продовжується і досягає критичного (порогового) рівня - такого значення МП, при якому різко збільшується деполяризація, - критичного потенціалу. Різниця між МП і критичним потенціалом називається пороговим потенціалом. При зменшенні МП на величину, що дорівнює пороговому потенціалу виникає потенціал дії (швидкі зміни МП, електричний імпульс). Він складається з фази деполяризації і реполяризації, які відповідають висхідній і низхідній кривій хвилі збудження. МП зменшується за абсолютною величиною до нуля і змінює свій знак на протилежний. Пік потенціалу дії припадає на період, коли відбувається перезарядка мембрани - реверсія потенціалу. Зовнішня сторона мембрани заряджується негативно, внутрішня - позитивно. Після цього починається фаза реполяризації - відновлення вихідного рівня поляризації. Проникність мембрани для іонів Nа+зменшується, а для К+ підвищується. Іони К+ дифундують з клітини на зовнішню поверхню мембрани, заряджуючи її позитивно. У період, коли проникність мембрани для К+ у ході реполяризація знижується, і реполяризація проходить повільніше, ніж у низхідній частині піку ПД, то спостерігається гіпополяризація мембрани (негативний слідовий потенціал). Відновлюється вихідна величина МП. Після цього у багатьох клітинах спостерігається ще деякий час підвищена проникність мембрани для К+, у зв’язку з цим МП починає зростати - відбувається гіперполяризація мембрани (виникає позитивний слідовий потенціал).
Генеруючи ПД клітина кожен раз отримує якусь кількість Nа+ і втрачає К+. Проте концентрація іонів у клітині і міжклітинній речовині не вирівнюється, що зумовлено дією натрієво-калієвої помпи, яка виводить Nа+ з клітини, і пропускає в клітину К+.
3. Під час процесу збудження змінюється збудливість тканин. Виділяють періоди збудливості:
1. Початкове зростання збудливості. Спостерігається під час місцевої (локальної) відповіді.
2. Рефрактерний - тимчасове зниження збудливості тканини. Розрізняють фази:
- абсолютної рефрактерності - повна незбудливість у період зростання ПД, збудження у цій фазі викликати неможливо, навіть якщо діє подразник надпорогової сили.
- відносна рефрактерність - знижена збудливість у період зменшення ПД, щоб викликати збудження необхідно подіяти подразником надпорогової сили.
2. Супернормальний - підвищеної збудливості, можна викликати збудження дуже слабким подразником підпорогової сили. Відповідає слідовому негативному потенціалу.
3. Субнормальний - пониженої збудливості порівняно з вихідним її рівнем. Зівпадає з позитивним слідовим потенціалом. Після чого відновлюється вихідний рівень збудливості.
Транспорт крізь мембрану органічних молекул та макромолекулярних комплексів.
Існує декілька механізмів транспорту речовин через мембрану.
Дифузія - проникнення речовин через мембрану по градієнту концентрації (з області, де їх концентрація вища, в область, де їх концентрація нижче). Дифузний транспорт речовин (води, іонів) здійснюється за участю білків мембрани, в яких є молекулярні пори, або за участю ліпідної фази (для жиророзчинних речовин).
При полегшеній дифузії спеціальні мембранні білки-переносники вибірково зв'язу
Активний транспорт зв'язаний з витратами енергії і служить для перенесення речовин проти їх градієнта концентрації. Він здійснюється спеціальними білками-переносниками, що утворюють так звані іонні насоси. Найбільш вивченим є Na-/ К- -насос в клітинах тварин, таких, що активно викачують іони Na назовні, поглинаючи при цьому іони К. Завдяки цьому в клітині підтримується велика концентрація К і менша Na в порівнянні з довкіллям. На цей процес витрачається енергія АТФ.
Взаємодія клітин, міжклітинні контакти.
Міжкліти́нні конта́кти — спеціалізовані ділянки поверхні клітини, якими вона кріпиться до інших клітин або позаклітинного матриксу[1]. До основних функцій міжклітинних контактів належить утримання клітин разом, забезпечення «спілкування» між ними, правильна їх орієнтація одна відносно іншої, регулювання переміщення клітин[2].
Злипання клітин між собою та приєднання їх до міжклітинного матриксу забезпечують молекули клітинної адгезії (англ. cell adhesion molecules, CAMs):кадгерини, інегрини, білки суперродини імунглобулінів, муцини та селектини. Цитоплазматична частина цих білків зазвичай приєднана до елементівцитоскелету. Більшість CAM рівномірно розподілені по поверхні цитоплазматичної мембрани і їх здебільшого достатньо для утримання клітин разом, проте в деякихтканинах на поверхні клітин формуються спеціалізовані ділянки скупчення молекул адгезії, саме для означення таких ділянок і використовують термін «міжклітинні контакти»
У залежності від того, чи контакти утворюються між однаковими чи різними клітинами, вони поділяються на гомо- та гетерофільні відповідно[3].
На основі структури і функцій клітинні контакти класифікують на чотири основні групи:
Якірні контакти (англ. anchoring junctions) — включають як з'єднання двох клітин, так і з'єднання клітин із позаклітинним матриксом завжди асоційовані з елементами цитоскелету: актиновими або проміжними філаментами. Не зважаючи на те, що якірні контакти мають відмінності у структурі, для них всіх характерна наявність трьох основних елементів: молекули клітинної адгезії (кадгеринів у випадку взаємодії клітина-клітина або інтегринів у випадку взаємодії клітина-позаклітинний матрикс), адаптерного білка, що кріпить CAM до цитоскелету, та пучка актинових або проміжних філаментів[3].
Адгезивні контакти (англ. adhesive junctions) або зони злипання (англ. zonula occludens) — з'єднують пучки актинових волокон у сусідніх клітинах. Сполучають латеральні поверхні епітеліальних клітин, навколо яких утворюють пояски, що розташовуються відразу ж під зоною щільних контактів[3][4].
Контакти із позаклітинним матриксом, приєднані до актинових філаментів;
Десмосоми або пластинки прикріплення — з'єднують пучки проміжних філаментів у сусідніх клітинах;
Гемідесмосоми — контакти клітини із позаклітинним матриксом, схожі за будовою до десмосом.
Замикальні контакти (англ. occluding junctions) — контакти, що «зшивають» клітини між собою, при цьому їхні мембрани максимально зближуються, внаслідок чого формуються непроникні або вибірково проникні щільні шари.
Щільні контакти (англ. tight junctions, zonula occludens) — замикальні контакти хребетних тварин;
Септовані контакти (англ. septate junctions) — замикальні контакти безхребетних тварин.
Контакти, що формують канали, або комунікативні контакти сполучають цитоплазму сусідніх клітин і допомагають інтегрувати їхній метаболізм:
Щілинні контакти або нексуси (англ. gap junctions) — комунікативні контакти тварин;
Плазмодесми — цитоплазматичні містки між рослинними клітинами.
Контакти, що передають сигнали, беруть участь у передачі інформації між клітинами через мембрани.
Хімічні синапси;
Імунологічні синапси;
Трансмембранні клітинні контакти ліганд-рецептор (наприклад Delta-Notch, ephrin-Eph). Будь-який із трьох інших типів клітинних контактів може виконувати крім структурної сигнальну функцію.
Неупорядковані, упорядковані утвори плазмолеми.
Вирости можуть бути неупорядковані (псевдоподії, мікроворсинки), упорядковані (посмугована облямівка), розміщені на вільній поверхні клітини і патологічні. Впинання звичайно бувають по базальному полюсу (у клітинах покручених канальців нирки). Клазматозом називають видалення окремих структурних компонентів клітини внаслідок відривання клітинних складок.
Структуровані вирости клітини: війки, джгутики.
Деякі клітини мають специфічні органели — війки та джгутики, які забезпечують рух, переважно, одноклітинних організмів. Ці струтури є у деяких клітин багатоклітинних організмів (війчастий епітелій). Війки та джгутики являють собою вирости цитоплазми, оточені клітинною мембраною. В середині виростів знаходяться мікротрубочки, скорочення яких надає клітині руху.
Війки - тонкі рухливі вирости поверхні клітин миготливого епітелію
Джгутики і війки мають вигляд тоненьких виростів цитоплазми діаметром приблизно 0,25 мкм. Вони вкриті плазматичною мембраною. Всередині цих органел розташована складна система з мікротрубочок. Джгутики і війки є у деяких одноклітинних організмів (хламідомонада, евглена, інфузорії), а також деяких типів клітин багатоклітинних (епітелій дихальних шляхів ссавців, сперматозоїди тварин, вищих спорових рослин тощо).
Р ухи
війок у цілому нагадують роботу
весел і, як правило, скоординовані
(наприклад, в інфузорій). Для джгутиків
характерний гвинтоподібний або
хвилеподібний рух (мал. 6).
ухи
війок у цілому нагадують роботу
весел і, як правило, скоординовані
(наприклад, в інфузорій). Для джгутиків
характерний гвинтоподібний або
хвилеподібний рух (мал. 6).
Мал. 6. Схема руху джгутика (1) і війки (2)
Джгутики і війки рухаються завдяки енергії, що вивільняється під час розщеплення молекул АТФ. Ці органели забезпечують не лише пересування клітин, а й надходження частинок їжі до них (наприклад, рух джгутиків травних клітин гідри). Вони можуть також виконувати чутливу (наприклад, у війчастих червів) і захисну (війки епітелію носової порожнини) функції.
Адгезивні. ізолюючи та комунікаційні міжклітинні контакти.
Межклеточние контакти творяться у місцях дотику клітин на тканинах і служать для міжклітинного транспорту речовин і передачі сигналів, і навіть для механічного скріплення клітин друг з одним. Класифікація міжклітинних контактів
1.Замикающіе міжклітинні контакти.
а) простий або пухкий контакт;
б) щільний замикає контакт.
2.Адгезіонние міжклітинні контакти.
а) точкові контакти;
б) адгезійні паски;
в) адгезійні з'єднання між клітиною і внутрішньоклітинним матриксом;
г) десмосоми.
3.Проводящіе.
а) нексус;
б) синапси.
Складні або адгезійні міжклітинні з'єднання представляють собою невеликі парні спеціалізовані ділянки плазматичних мембран двох сусідніх клітин. Вони поділяються на замикаючі (ізолюючі), зчіплюють (заякориваются) і комунікаційні (об'єднуючі) контакти.
До замикаючим (ізолюючим) відноситься щільний контакт (замикає зона - zona occuludens). У цьому з'єднанні беруть участь спеціальні інтегральні білки, розташовані на поверхні сусідніх клітин, що утворюють подобу комірчастої мережі. Ця чарункова мережаоточує у вигляді паска весь периметр клітини, з'єднуючись з такою ж мережею на поверхні сусідніх клітин. Ця область непроникна для макромолекул та іонів і, отже, вона замикає, відмежовує міжклітинні щілини (і разом з ними власне внутрішнє середовище організму) від зовнішнього середовища. Цей тип з'єднань характерний для клітин одношарових епітеліїв та ендотелію.
До зчіплюються, або заякориваются, з'єднань відносяться адгезивний (Сцепляющій) пасок і десмосоми. Спільним для цієї групи сполук є те, що до ділянок плазматичних мембран з боку ци-топлазми підходять фібрилярні елементи цитоскелету, які як би заякориваются на їх поверхні.
Адгезивний (Сцепляющій) пасок - парне утворення у вигляді стрічки, що оперізує апикальную частина клітини одношарових епітеліїв. Тут клітини пов'язані один з одним інтегральними гликопротеидами, до яких з боку цитоплазми і тієї й іншої клітки примикає шар примембранних білків, що включають характерний білок вінкулін. До цього шару підходить і зв'язується з ним пучок Актинові мікрофіламентів. Кооперативний скорочення Актинові мікрофіламентів в багатьох сусідніх клітинах може призвести до зміни рельєфу всього епітеліального пласта.
До зчіплюють з'єднанням може бути віднесений так званий фокальний контакт, характерний для фібробластів. У цьому випадку клітина з'єднується не з сусідньою кліткою, а з елементами позаклітинного субстрату. В освіті фокальної контакту також беруть участь актинові мікрофіламенти. До заякориваются міжклітинним сполук відносяться і десмосоми. Це теж парні структури, що представляють собою невеликий майданчик або біла пляма діаметром близько 0,5 мкм. З боку цитоплазми до плазматичної мембрани прилягає шар білків, до складу якого входять десмоплакіни. У цьому шарі якоря пучки цитоплазматичних проміжних філаментів. Із зовнішнього боку плазмолеми сусідніх клітин в області десмосом з'єднуються за допомогою трансмембранних доменів білків - десмоглеінов. Кожна клітина епідермісу шкіри може мати до кількох сотень десмосом.
Функціональна роль десмосом полягає головним чином у механічного зв'язку між клітинами. Десмосоми зв'язують один з одним клітини в різних епітелію, в сердечних і гладких м'язах. Полудесмосомами пов'язують епітеліальні клітини з базальною мембраною.
Комунікаційні з'єднання в клітинах тварин представлені так званими щілинними контактами і синапсами.
Щілинне з'єднання, або нексус, являє собою область протяжністю 0,5-3 мкм, де плазмолеми розділені проміжком в 2-3 нм. З боку цитоплазми ніяких спеціальних примембранних структур у цій області не виявляється, але в структурі плазмолемма сусідніх клітин один проти одного розташовуються спеціальні білкові комплекси (коннексони), які утворюють як би канали з однієї клітини в іншу. Цей тип з'єднання зустрічається у всіх групах тканин.
Функціональна роль щілинного з'єднання полягає в перенесенні іонів і дрібних молекул (молекулярна маса 2 -10) від клітини до клітини. Так, у серцевому м'язі збудження, в основі якого лежить процес зміни іонної проникності, передасться від клітини до клітини через нексус.
Гіалоплазма клітини. її хімічний склад та функції
Гіалоплазма (від грец. hyalos — скло і plasma — виліплене) (синоніми: цитоплазматичний матрикс (від лат. matrix — мати), основна речовина цитоплазми чи цитозоль) — це організована, впорядкована багатокомпонентна система, яка здатна до формування складних структур. У гіалоплазмі знаходяться всі будівельні компоненти, необхідні для утворення мембран, мікрофіламентів, мікротубул, гранул. До складу цитоплазми входять також нуклеотиди: аденозинтрифосфат (АТФ), гуанозинтрифосфат (ГТФ), тимідинтрифосфат (ТТФ), цитозинтрифосфат (ЦТФ) і уридинтрифосфат (УТФ). У цитоплазмі важливі життєві процеси здійснюють ферменти, які беруть участь у проміжному обміні клітин (наприклад, гліколітичні ферменти). У ній знаходяться також інші, малі молекули та іони, які мають важливе значення для клітинних функцій та для підтримування внутрішньоклітинного і міжклітинного середовища.
Треба відзначити, що роль гіалоплазми в клітині ще слабо вивчена. Вважають, що вона виконує такі основні функції: (1) інтегральну — об'єднання всіх клітинних структур і забезпечення їх хімічної взаємодії; (2) гліколіз — анаеробне (безкисневе) розщеплення вуглеводів; (3) функціонування ферментних сигнальних систем (наприклад, проліпазних) та (4) транспорт речовин. Гіалоплазма — це місце більшої частини реакцій проміжного обміну, синтезу білка на вільних полісомах, синтезу жирних кислот.
Гіалоплазма або матрикс цитоплазми становить внутрішнє середовище клітини. Вона складається з води (90 %) і різних біополімерів (7 %) білків, нуклеїдових кислот, полісахаридів, ліпідів, з яких основну частину становлять білки різної хімічної й функціональної специфічності. У гіалоплазмі втримуються також амінокислоти, моносахару, нуклеотиди й інші низькомолекулярні речовини. Бозна-чиї з'єднання утворять із водою колоїдну систему, що залежно від умов може бути більше щільної (у формі гелю) або більше рідкої (у формі золю) як у всій цитоплазмі, так і в окремих її ділянках. У гіалоплазмі локалізуються й взаємодіють між собою й середовищем гіалоплазми різних органелл й включення. При цьому розташування їх найчастіше специфічно для певних типів клітин.
Через біліпідну мембрану Гіалоплазма взаємодіє з позаклітинним середовищем. Отже, Гіалоплазма є досить динамічним середовищем і відіграє важливу роль у функціонуванні окремих органелл і життєдіяльності клітки в цілому.
Компартментаиія клітини.
компартментація – це просторове озмежування клітини внутрішніми мамбранами на відсіки, що дає змогу здійснювати перебіг багатьох біохімічних реакцій одночасно й незалежно одна від одної; кожен відсік має у своєму складі специфічні білки, що визначають його унікальні функції; Компартментація - розмежування фонду іонів і низькомолекулярних сполук у живій клітині на окремі ділянки (компартменти), що відрізняються функціональним значенням і інтенсивністю участі в обміні речовин. Здійснюється за участю мембран і органелклітини. Компартментація має велике значення для внутрішньоклітинної організації, обміну речовин і його регуляції
Найважливіша функція: сприяє компартментаціі - підрозділу вмісту клітини на окремі осередки, що відрізняються деталями хімічного або ферментного складу. Цим досягається висока впорядкованість внутрішнього вмісту будь еукаріотичної клітини. Компартментація сприяє просторовому розділенню процесів, що протікають в клітині. Окремий компартмент (осередок) представлений будь-якої мембранної органели (наприклад, лізосомах) або її частиною (кристами, відмежованими внутрішньою мембраною мітохондрій).
Структура та функція мембраннпх органів
Кожа человека - это барьер, защищающий клетки, её мы и называем –клеточной мембраной. Она не позволяет компонентам клетки (цитоплазме) вытечь наружу. Главная задача клеточной мембраны - это удерживать клетку в целостности, при этом определять, что может попасть внутрь клетки, а что может оттуда выйти. Клетки любого организма имеют клеточные мембраны, даже клетки бактерий.
Строение клеточной мембраны

Состоит клеточная мембрана из бинарного ряда липидов. Располагаются молекулы липидов в два ряда и каждый ряд точно такой же, как предыдущий. Структуру молекулы липида - эти две части единого целого, как раз и отображают. Ещё эти две части единого целого называют – гидрофобной (водонепроницаемой) и гидрофильной секциями.
Гидрофобная секция не любит воду и подобных воде молекул, благодаря бинарному слою липидов выступает вроде защитного механизма.
Гидрофильная секция напротив способна притягивать воду и подобные воде молекулы, после чего выталкивает их наружу. В итоге получается такая базовая жидкая мозаичная модель.
Жидкостно-мозаичная модель
Открыли жидкую мозаичную модель клеточной мембраны в 1972 году. Эта модель демонстрирует структуру размещения протеинов внутри или на биполярном слое липидов. Размещаются протеины в хаотичном порядке, при этом получается мозаика протеинов. Протеины пересекающие бинарный ряд липидов, играют важную роль в транспортировке маленьких молекул через мембрану.
Функции клеточной мембраны
В результате того, что клеточная мембрана имеет среду полупроницаемую, то только некоторые виды самых мелких молекул способны проникнуть внутрь и наружу клетки сквозь мембрану. Называется данный процесс – диффузией. Но для того, чтобы такое действие произошло, мембрана должна быть вроде открытых дверей, то есть проницаемой, чтобы маленькая молекула смогла проникнуть сквозь неё. В том случае, когда мембрана непроницаема, маленькая молекула не имеет возможности проникнуть, здесь можно сказать «двери закрыты». Однако следует учесть, что молекула может переместиться только из переполненной области, в более свободную. Например, аминокислота желает пробраться через мембрану к самой клетке, при этом клеточная мембрана открыта для неё, это может произойти в том случае если концентрация аминокислоты за пределами клетки больше, чем в самой клетки. Только при таких условиях произойдёт процесс диффузии.
Следующий процесс, в котором главную роль играет клеточная мембрана - это так называемая помощь при транспортировке. Некоторые маленькие молекулы в мембране которых разместились протеины, способны помочь при пересечении мембраны. Вроде протеинового эскорта сквозь неё. Данный процесс напоминает процесс диффузии, так как протеиновая помощь при пересечении мембраны происходит, когда молекулы переходят из области высокой концентрации, с целью понизить её. Или же просто из области высокой концентрации молекул в область менее загруженную.
Опять же, молекулы способны пересекать клеточную мембрану способом активной транспортировки. Такой вид транспортировки представляет собой переход молекул из области с низкой концентрации в противоположную область с высокой концентрацией. Такой процесс транспортировки противоположный диффузии и посреднической транспортировки, осуществляется против правил концентрационной направленности. Для того чтобы такие передвижения осуществлять необходима концентрация энергии в форме аденозинтрифосфата (АТФ). АТФ- это нуклеотид, который состоит из аденина, рибозы и трёх остатков фосфорной кислоты. Он является универсальным накопителем и переносчиком химической энергии, известным для всех организмов и клеток. АТФ применяется для некоторых клеточных действий, начиная движениями и заканчивая репродукцией.
Гранулярна і та агранулярна ендоплазматична сітка га її функції.
Ендоплазматична сітка (від грец. endos — внутрішній), або ендоплазматичний ретикулум (від лат. reticulum — сітка) — це мембранна органела, яка ділить цитоплазму на компартменти (від англ. сompartment — відділ, відсік). Ендоплазматична сітка — це порожниста система у вигляді замкненої сукупності канальців і цистерн, утворених суцільною безперервною мембраною і заповнена матриксом. Матрикс (від лат. matrix — мати) — це пухкий матеріал помірної щільності (продукт синтезу). Ендоплазматична сітка відкривається в перинуклеарний простір — простір між двома мембранами каріолеми. Це — синтетичний і частково транспортний апарат цитоплазми, що забезпечує виконання клітиною складних функцій.
Біохімічний склад мембран ендоплазматичної сітки, виявлений шляхом диференціального центрифугування, показав, що сітка містить білки і ліпіди, серед яких багато фосфоліпідів, а також ферменти: аденозинтрифосфатазу, ферменти синтезу мембранних ліпідів. При виготовленні мікроскопічних препаратів цитоплазма клітин з добре розвиненою гранулярною ендоплазматичною сіткою фарбується основними барвниками. У класичній гістології базофільна субстанція (тигроїд, або субстанція Нісля), описана в нейронах, є скупченням угрупувань гранулярної ендоплазматичної сітки.
Розрізняють два види ендоплазматичної сітки: гладку (агранулярну, аЕС), представлену трубочками, що анастомозують між собою, ішорстку (гранулярну, грЕС), побудовану з цистерн, також з’єднаних між собою і вкритих полісомами. Деякі автори виділяють ще перехідну, або транзиторну (тЕС) ендоплазматичну сітку, яка знаходиться в ділянці переходу однієї різновидності ЕС в іншу.
Гранулярна ЕС присутня у всіх клітинах (за винятком сперматозоїдів), однак ступінь її розвитку різний і залежить від спеціалізації клітин. Так, сильно розвинену грЕС мають епітеліальні залозисті клітини (підшлункової залози, які виробляють травні ферменти, печінкові — синтезують альбуміни сироватки крові), фібробласти (клітини сполучної тканини, які продукують білок калоген), плазматичні клітини (продукують імуноглобуліни).
Агранулярна ЕС переважає в клітинах надниркових залоз (здійснює синтез стероїдних гормонів), в м’язових клітинах (бере участь в обміні кальцію), в клітинах фундальних залоз шлунка (участь у виділенні Сl–).
Функції ендоплазматичної сітки. Гладка (агранулярна) і шорстка (гранулярна) ендоплазматичні сітки своїми мембранами і вмістом (матриксом) виконують чотири спільні функції:
1) розмежувальну, структуруючу, яка забезпечує упорядкований розподіл цитоплазми, не допускає її змішування та потрапляння в органелу випадкових речовин;
2) трансмембранний транспорт, завдяки якому здійснюється перенесення через стінку мембрани необхідних речовин;
3) синтез мембранних ліпідів ферментами, які містяться в самій мембрані і забезпечують репродукцію ендоплазматичної сітки;
4) вважають, що різниця потенціалів, яка виникає між двома поверхнями мембран органели можливо забезпечує проведення імпульсів збудження.
Крім того, кожна з різновидностей сітки виконує свої спеціальні функції.
Агранулярна ендоплазматична сітка, крім названих функцій, спільних для обох видів ЕС, виконує ще й властиві лише їй функції:
1) Депо кальцію. У багатьох клітинах (скелетних і серцевих м’язах, нейронах, яйцеклітині) існують механізми, які забезпечують зміну концентрації Са2+, такі як Са2+-помпа, Са2+-канал, Са2+-зв’язувальний білок. У поперечнопосмугованій м'язовій тканині існує спеціалізована гладка ендоплазматична сітка, названа саркоплазматичним ретикулумом, яка є резервуаром Са2+, а її мембрани містять потужні кальцієві помпи, які в соті частки секунди можуть викидати велику кількість кальцію в цитоплазму або транспортувати його у порожнини каналів ретикулума (детальніше описана в курсі гістології в розділі про поперечносмугасту м'язову тканину).
2) Синтез ліпідів, речовин типу холестерину і стероїдних гормонів. Синтез стероїдних гормонів відбувається головним чином в ендокринних клітинах гонад і надниркових залоз, а також в клітинах печінки, нирок. У клітинах кишечника синтезуються ліпіди, які у вигляді хіломікронів виводяться в лімфу, а потім у кров.
3) Детоксикуюча функція — знешкодження екзогенних і ендогенних токсинів. Наприклад, в печінкових клітинах (гепатоцитах) є ферменти оксидази, які руйнують фенобарбітал.
4) Ферменти органели беруть участь у синтезі глікогену (у печінкових клітинах).
5) Дещо видозмінена гладка ендоплазматична сітка добре розвинена в парієтальних клітинах фундальних залоз шлунка. Там вона бере участь у механізмі, який забезпечує підтримання концентрації іонів хлору, необхідних для вироблення соляної кислоти як складової шлункового соку.
Гранулярна ендоплазматична сітка, крім названих спільних, виконує ще спеціальні функції.
1) Синтез білків на грЕС має деякі особливості . Починається він на вільних полісомах, які в дальшому зв’язуються з мембранами ЕС. На першому етапі взаємодії іРНК з рибосомами відбувається утворення особливого сигнального пептида, який контактує з сигнал-розпізнавальною частиною (СРЧ). Комплекс СРЧ-рибосома зв’язується з рецептором на мембрані ЕС, який називається причальним білком. Крім того на мембрані грЕС є інтегральні рецепторні білки рибофорини (зв’язані з порою), що забезпечують прикріплення великої субодиниці, а тим самим і всієї рРНК. Після цього сигнал-розпізнавальна частина відділяється від рибосоми і на рибосомі починається синтез білка. Під дією сигнальної пептидази від’єднується сигнальний пептид і синтез білка продовжується, а білок через пору в мембрані ЕС виділяється в її просвіт. Після закінчення синтезу субодиниці рибосом дисоціюють, звільнюються від іРНК і можуть знов включатися в процес синтезу.
Агранулярна ендоплазматична сітка, крім названих функцій, спільних для обох видів ЕС, виконує ще й властиві лише їй функції, зокрема:
— депо кальцію. У багатьох клітинах (скелетних і серцевих м’язах, нейронах, яйцеклітині) існують механізми, які забезпечують зміну концентрації Ca2+, такі як Ca2+ -помпа, Ca2+ – канал, Ca2+ – зв’язувальний білок. У поперечно-смугастій м’язовій тканині існує спеціалізована гладка ендоплазматична сітка,названа саркоплазматичним ретикулумом, яка є резервуаром Ca2+. Її мембрани містять потужні кальцієві помпи, які дуже швидко, протягом сотих часток секунди можуть викидати велику кількість кальцію в цитоплазму або транспортувати його у порожнини каналів ретикулума;
— синтез ліпідів, речовин типу холестерину і стероїдних гормонів. Синтез стероїдних гормонів відбувається головним чином в ендокринних клітинах гонад і надниркових залоз, а також у клітинах печінки, нирок. У клітинах кишечника синтезуються ліпіди, які у вигляді хіло- мікронів виводяться в лімфу, а пол’м у кров;
— детоксикуюча функція — знешкодження екзогенних і ендогенних токсинів. Наприклад, в печінкових клітинах (гепатоцитах) містяться ферменти оксидази, які руйнують фенобарбітал;
— ферменти органели беруть участь у синтезі глікогену (у печінкових клітинах);
— дещо видозмінена гладка ендоплазматична сітка добре розвинена в парієтальних клітинах фундальних залоз шлунка.
Особливості будови комплексу Гольджи та його функції.
Комплекс Ґольджі (також званий як Апарат Ґольджі, тільце Ґольджі та інші) —одномембранна органела, що є переважно в еукаріотів.
Була відкрита у 1898 році італійським лікарем Камілом Ґольджі і була названа в честь нього. Основна функція комплексу Ґольджі — це гліколізація та фосфоризація речовин зендоплазматичного ретикулуму. Це система паралельно розташованих та сплющених цистерн і трубочок, до яких прикріплюються мембранні міхурці, що транспортують речовини від ендоплазматичної сітки.
Ця мембранна органела представлена трьома видами утворів: дископодібними мембранними мішечками (цистернами), розміщеними пучками щільно на відстані 14-25 нм з внутрішнім простором 5-20 нм (частіше по 5-6 мішечків у комплексі); системою трубочок діаметром 20-50 нм; і міхурців різних розмірів. Мішечки сполучаються між собою і мають трубочкове з'єднання з іншими такими ж апаратами. У рослинних клітинах виявляється ряд окремих стопок, який називають диктіосомою. Диктіосоми можуть бути відділені одна від одної прошарками цитоплазми або з'єднаними у комплекс. В тваринних клітинах часто міститься одна велика або кілька з'єднаних трубками стопок.
Основною структурною одиницею комплексу Гольджі* є купка плоских цистерн (мішечків), оточених мембраною. Поруч із цистернами розташовані пухирці та канальці. Цистерни комплексу Гольджі, як правило, полярні: тобто, до одного з полюсів постійно підходять пухирці, які відриваються від ендоплазматичної сітки. Вони містять речовини, які там утворилися. Зливаючись із цистернами комплексу Гольджі, пухирці віддають їм свій вміст. Одночасно від другого полюса цистерн відходять пухирці з різними речовинами, завдяки чому вони транспортуються в інші ділянки клітини та виводяться з неї (мал. 5).

Мал. 5. Комплекс Гольджі:
1 - трубочки;
2 - пухирці;
3 - стопки цистерн
Комплекс Гольджі є в усіх еукаріотичних клітинах, але у різних організмів його будова може відрізнятись. Так, у клітинах рослин структурні компоненти цієї органели (їхня кількість може сягати 20 і більше) відокремлені один від одного, а в клітинах інших організмів вони зазвичай з'єднані.
Лізосоми. види лізосом.
Лізосоми (від грецьк. lysis — розчинення і soma — тіло) — полімерні мембранні органели, які містяться в клітинах майже всіх типів. Іншими словами, лізосоми — це органели, які забезпечують катаболіч- ні процеси в потрібному місці у потрібний час.
Будова лізосом. Лізосоми мають вигляд міхурців діаметром приблизно 0,5 мкм, оточених мембраною і заповнених гідролітичними ферментами, що діють у кислому середовищі (кислі гідролази). Ферментний склад лізосом Дуже різноманітний, вони здатні розщеплювати біополімери різного хімічного складу: білки (гідроліз забезпечують протеолітичні ферменти), вуглеводи (гліколітичні ферменти), ліпіди (ліполітичні ферменти). У лізосомах виявлено також ендонуклеази, фосфоліпази, деякі фосфатази і сульфатази. У неробочому стані ферменти неактивні: 80 % їх інактивовані глікозаміногліканами вмісту міхурців і 20 % — мембранами.
Функції лізосом — це аутоліз (від грецьк. autos — сам і lysis — розчинення) і гетероліз (від грецьк. heteros — інший). Розрізняють фізіологічний аутоліз (розсмоктування хвоста пуголовка, підгрудинної залози у підлітків) і патологічний, наприклад, лізис клітин печінки при отруєнні). Гетероліз є розщепленням чужих речовин — ксенобіотиків (наприклад, при фагоцитозі чи піноцитозі). Умовами функціонування лізосом є: 1) наявність рецепторів, здатних сприймати фагосоми чи піноцитозні міхурці або відпрацьовані структури клітини; 2) переміщення лізосоми по цитоплазмі, яке відбувається за участю мікро- тубул; 3) здатність руйнувати мембрани в місці контакту лізосоми з фагосомою (фагоцитованою частинкою) або піноцитозним міхурцем. Види лізосом. Останнім часом класифікація лізосом уточнена. Так, міхурці, заповнені гідролітичними ферментами, яких раніше вважали первинними лізосомами, тепер називають пізніми (перинуклеарними) ендосомами. Функції лізосом зумовлюються їх будовою, і вчені тому виділяють різні види лізосом, які можна виявити при електронно- мікроскопічному дослідженні.
Види
В останній час класифікація лізосом уточнена. Так, міхурці, заповнені гідролітичними ферментами, яких раніше вважали первинними лізосомами, тепер називають пізніми (перинуклеарними) ендосомами. Функції лізосом відкладають відбиток на їх будову і тому виділяють різні види лізосом, які можна виявити при електронномікроскопічному дослідженні.
Фаголізосоми (фаголізоми), або гетерофагосоми (від грец. heteros — інший, phagein — пожирати і soma — тіло) утворюється шляхом поєднання пізніх ендосом або лізосом з фагосомами або піноцитозними міхурцями, які містять захоплений клітиною матеріал для внутрішньоклітинного перетравлювання. Активні ферменти в них безпосередньо контактують з біополімерами, які підлягають розщепленню. Процес розщеплення цих полімерів називається гетерофагією.
Аутофаголізосоми утворюються при злитті пізньої ендосоми або лізосоми з аутофагосомою (від грец. autos — сам, phagein —пожирати і soma — тіло), тобто міхурцем, який містить власні макромолекулярні комплекси клітини, наприклад, цілі клітинні органели, або їх фрагменти, які втратили функціональні здатності і підлягають дезінтеграції. Процес розщеплення цього матеріалу називаютьаутофагією.
Мультивезикулярними тільцями (від лат. multi — багато і vesicula — міхурець) називають вакуолі з великою кількістю міхурців. Вони утворюються шляхом злиття ранніх ендосом з пізніми. Наявні в органелі ферменти забезпечують поступове руйнування внутрішніх міхурців.
Залишкові тільця — це оточені мембраною нерозщеплені частинки, що можуть тривалий час залишатися в цитоплазмі і тут утилізуватися або шляхом екзоцитозу виводитися поза клітину. У залишкових тільцях нагромаджується матеріал, розщеплення якого ускладнено; найчастіше це ендогенний пігмент коричневого кольору — ліпофусцин (“пігмент старіння” чи “зношування”).
Мієлінові фігури є автофагосомами з нагромадженим мембранним матеріалом, часто щільно концентрично упакованим, що довго залишається в клітині.
Недостатність того чи іншого фермента в лізосомі веде до скупчення нерозщеплених речовин в таких кількостях, що порушуються функції клітин, а це призводить до лізосомальних хвороб нагромадження.
Пероксисоми.
Пероксисоми (мікротільця) є органелами у вигляді міхурців діаметром 0,05–1,5 мкм, оточених мембраною і заповнених дрібнозернистим матриксом, що у центрі (серцевині) містить волокнисті та трубчасті структури і щільний кристалоїд. У пероксисомах виявлено ферментні системи, склад яких може дещо змінюватися. Основними з них є ферменти окиснення амінокислот та перекисного окиснення — каталаза і пероксидаза, оксидаза d-амінокислот і уратоксидаза. Серцевина відповідає ділянці конденсації ферментів.
Утворення пероксисом відбувається шляхом відбруньковування їх від агранулярної ЕС, ферменти їх синтезуються частково в гранулярній ЕС, а частково — в гіалоплазмі. Вважають, що нові пероксисоми утворюються шляхом розщеплення існуючих завдяки постійному надходженню ферментів, а також завдяки відокремленню нових пероксисом після збільшення розмірів існуючих унаслідок збагачення їх ферментами, що поступають з гіалоплазми. Пероксисоми оновлюються кожних 5-6 днів.
Функції пероксисом. Цим органелам належить важлива роль у процесах внутрішньоклітинної детоксикації. Каталаза розщеплює пероксид водню (Н2О2), який утворюється в процесах перекисного окиснення і є отруйним для клітин. Ферменти пероксисом забезпечують також розщеплення сечової кислоти, беруть участь в ряді катаболічних і анаболічних реакцій, в обміні амінокислот, поліамінів, оксалату, у регуліції обміну ліпідів. У пероксисомах печінкових клітин розщеплюється до 50% поглинутого етилового спирту.
Мітохондрії.
М ітохондрія
(від грец.
μιτος або mitos — «нитка» та κουδριον або
khondrion — «гранула») — двомембранна
органела,
присутня у більшості клітин
еукаріот.
Мітохондрії іноді називають «клітинними
електростанціями», тому що вони
перетворюють молекули поживних речовин
на енергію у формі АТФ
через процес відомий як окислювальне
фосфорилювання. Типова еукаріотична
клітина містить близько 2 тис. мітохондрій,
які займають приблизно одну п'яту її
повного об'єму. Мітохондрії містять так
звану мітохондріальну
ДНК, незалежну від ДНК,
розташованої у ядрі
клітини. Відповідно до загальноприйнятої
ендосимбіотичної
теорії, мітохондрії походять з
вільноживучих клітин прокаріот,
родичів сучасних протеобактерій.
ітохондрія
(від грец.
μιτος або mitos — «нитка» та κουδριον або
khondrion — «гранула») — двомембранна
органела,
присутня у більшості клітин
еукаріот.
Мітохондрії іноді називають «клітинними
електростанціями», тому що вони
перетворюють молекули поживних речовин
на енергію у формі АТФ
через процес відомий як окислювальне
фосфорилювання. Типова еукаріотична
клітина містить близько 2 тис. мітохондрій,
які займають приблизно одну п'яту її
повного об'єму. Мітохондрії містять так
звану мітохондріальну
ДНК, незалежну від ДНК,
розташованої у ядрі
клітини. Відповідно до загальноприйнятої
ендосимбіотичної
теорії, мітохондрії походять з
вільноживучих клітин прокаріот,
родичів сучасних протеобактерій.
Мітохондрія оточена внутрішньою і зовнішньою мембранами, складеними з подвійного шару фосфоліпідів і білків. Ці дві мембрани схожі, проте, мають різні властивості. Зовнішня мембрана гладенька, вона не утворює ніяких складок і виростів, товщиною 7 нм. Внутрішня мембрана утворює численні складки, спрямовані в порожнину мітохондрії, товщиною 7 нм. Через цю двомембранну організацію мітохондрія фізично розділена на 5 відділів. Це зовнішня мембрана, міжмембранний простір (простір між зовнішньою і внутрішньою мембранами, 10 нм), внутрішня мембрана, криста (сформована складками внутрішньої мембрани) і матрикс (простір в межах внутрішньої мембрани). Мітохондрія має від 1 до 10 мікрон (μм) за розміром. Зовнішня мембрана
Зовнішня мітохондріальна мембрана, що оточує всю органелу, має співвідношення фосфоліпідів до білків подібне до плазматичної мембрани еукаріот (близько 1:1 за вагою). Вона містить численні інтегральні білки - поріни, які мають відносно великий внутрішній канал (близько 2-3 нм), що пропускає всі молекул від 5000 Да та менше. Більші молекули можуть перетнути зовнішню мембрану тільки за допомогою активного транспорту. Зовнішня мембрана також містить ферменти, залучений в такі різноманітні активності як подовження жирних кислот, окислення адреналіну і біодеградація триптофану.
Міжмембранний простір
Міжмембранний простір – це простір між зовнішньою та внутрішньою мембраною мітохондрії. Його товщина становить близько 10-20 нм. Оскільки зовнішня мембрана мітохондрії проникна для невеликих молекул та іонів, їх концентрація в периплазматичному просторі мало відрізняється від їхньої концентрації в цитоплазмі. Для транспортування великих білків, навпаки, необхідні специфічні сигнальні пептиди; тому білкові компоненти периплазматичного простору та цитоплазми відрізняються. Одним из білків, що містяться у периплазматичному просторі, є цитохром c — один з компонентів дихального ланцюга мітохондрій.
Внутрішня мембрана
Зображення кріст в мітохондрії печінки пацюка
Внутрішня мітохондріальна мембрана містить білки з чотирма видами функцій [2]:
Білки, що проводять окислювальні реакції респіраторного ланцюжка.
АТФ синтаза, яка виробляє в матриці АТФ.
Специфічні транспортні білки, які регулюють проходження метаболітів між матріцею і цитополазмою.
Системи імпорту білків.
Внутрішня мембрана містить більш ніж 100 різних поліпептидів і має дуже високе співвідношення фосфоліпідів до білків (більш ніж 3:1 за вагою, тобто приблизно 1 білок на 15 молекул фосфоліпідів). Додатково, внутрішня мембрана багата на незвичайний фосфоліпід кардіоліпін, який є зазвичай характеристикоюбактерійних плазматичних мембран. На відміну від зовнішньої мембрани, внутрішня мембрана не містить порінів і тому надзвичайно непроникна; майже всі іони і молекули вимагають спеціальних мембранніих транспортних білків для проникнення до та з матриксу. Крім того, через внутрішню мембрану підтримується мембранний потенціал.
Внутрішня мембрана розділяється на численні кристи, які розширюють зовнішню область внутрішньої мітохондріальної мембрани, збільшуючи її здатність виробляти АТФ. У типової мітохондрії печінки, наприклад, зовнішня область, зокрема кристи, — приблизно у п'ять разів перевищує площу зовнішньої мембрани. Мітохондрії клітин які мають вищі потреби в АТФ, наприклад, м'язові клітини, містять більше крист, ніж типова мітохондрія печінки.
Мітохондріальний матрикс
Матрикс — простір, обмежений внутрішньою мембраною. Матрикс містить надзвичайно сконцентровану суміш сотень ферментів, на додаток до спеціальних мітохондріальних рибосом, тРНК і декількох копій мітохонднальноїної ДНК. Головні функції ферментів включають окислення пірувата і жирних кислот, та лимонно-кислотний цикл.
Мітохондрії мають свій власний генетичний матеріал і системи для виробництва власної РНК і білків. Ця нехромосомна ДНК кодує нечисленні мітохондріальні пептиди (13 у людини), що використовуються у внутрішній мітохондріальній мембрані разом з білками що кодуються генами клітинного ядра.
Гіпотеза симбіотичного походження еукаріотичної клітини.
Сценарії симбіотичного походження евкаріотичної клітини: гіпотеза двох плазм (Мережковський, 1905), гіпотеза хімеризації (Zillig, 1989), воднева гіпотеза (Martin, Muller, 1998).
Еволюційних змін у будові прокаріотів майже не відбувалось: усі їхні викопні форми не відрізняються від сучасних. Це пояснюють насамперед тим, що серед них немає багатоклітинних організмів, тобто прокаріоти нездатні до диференціації клітин та ускладнення організації. Внаслідок простої будови геному (єдина кільцева молекула ДНК) у них легко відбувається горизонтальний (між різними видами) транспорт спадковоїінформації за допомогою переносу фрагментів ДНК вірусами-бактеріофагами з однієї бактеріальної клітини в іншу. Крім того, спадкові властивості бактеріальних клітин можуть значно змінюватися під впливом плазмід.
Плазміди, або позаклітинні фактори спадковості, - ланцюжки з кількох нуклеотидів. Вони часто перебувають поза організмами і за певних умов можуть потрапляти всередину прокаріотних клітин і вбудовуватися в їхні ДНК. Тому у прокаріотів відбувається здебільшого біохімічна, або функціональна, еволюція. Вони здатні опановувати різні адаптаційні зони, виробляючи нові ферменти для засвоєння тих чи інших поживних речовин.
В еукаріотів подібні форми зміни спадковості через складність геному та взаємодію генів призводять до загибелі клітини і тому не можуть бути факторами еволюції.
У прокаріотних екосистемах відсутні ланцюги виїдання через нездатність бактерій до фагоцитозу. Тому ланцюги живлення прокаріотних біогеоценозів мало розгалужені, а їхня видова різноманітність незначна. Отже, прокаріотні екосистеми мають невисоку здатність до саморегуляції.
Протягом першої половини протерозойської ери (почалась 2,5 млрд, закінчилась - близько 0,6 млрд років тому) прокаріотні екосистеми опанували весь світовий океан. Близько 2 млрд років тому з'явилися первісні одноклітинні еукаріоти, які швидко дивергували на рослини (водорості), тварин (найпростіші) та гриби.
Наприкінці ХІХ ст. видатний ботанік та фізіолог рослин М. С. Цвєт, на основі спостережень поділу хлоропласту та ядерця у зеленої водорості Spirogyra висловив припущення, що клітини рослин можливо представляють собою симбіотичний утвір з клітини-господаря та бактеріальних внутрішньоклітинних симбіонтів, які відомі як хлоропласти та ядерця. Це припущення у 20-40-х роках ХХ ст. було детально розроблено, в першу чергу, К. С. Мережковським у так звану ендосимбіотичну гіпотезу походження евкаріотичної клітини. Проте ідеї К. С. Мережковського не знайшли підтримки у широкого загалу біологів. Гіпотезу ендосимбіотичного походження евкаріот називали вражаючим витвором людської фантазії", «луна-парком», «непристойною темою у колі освічених біологів»[2]. У 60-х роках гіпотеза ендосимбіотичного походження евкаріотичної клітини була заново сформульована американською дослідницею Л. Маргеліс.
Згідно з гіпотезою Мережковського-Маргеліс, евкаріотична клітина виникла внаслідок кількох ендосимбіозів: гіпотетична прокаріотична анаеробна клітина, здатна до фагоцитозу, захопила, проте не перетравила, а зберегла в цитоплазмі, аеробну гетеротрофну бактерію, яка трансформувалась в мітохондрію. Далі клітина-господар, що містила мітохондрію, вступила в симбіоз з рухливою спірохетоподібною гетеротрофною бактерією, яка дала початок джгутику. Після цього внаслідок автогенетичного процесу, пов'язаного з утворенням глибоких інвагінацій плазмалеми, навколо нуклеоїду клітини-господаря утворилось ядро. Далі базальні тіла джгутиків трансформувались в центри організації веретена поділу, внаслідок чого виник спочатку мітоз, а потім його модифікація — мейоз, разом із статевим процесом. Виникла перша гетеротрофна евкаріотична клітина. Нащадки цієї клітини, еволюція яких пішла по шляху вдосконалення фагоцитозу, дали початок тваринам. Ті ж нащадки, які еволюціонували в напрямку удосконалення осмотрофного живлення, дали початок грибам. Симбіоз гетеротрофної евкаріотичної клітини з синьозеленою водорістю призвів до появи рослин, оскільки симбіотична синьозелена водорість надалі трансформувалась у пластиду.
Як спосіб досягнення біологічного прогресу для еукаріотів характерне ускладнення організації в процесі історичного розвитку. Вже в одноклітинних організмів (водорості, інфузорії) клітини побудовані дуже складно. Поява багатоклітинних організмів - ще один прояв здатності еукаріотів до ускладнення будови. Більшість учених вважає, що багатоклітинні організми походять від колоніальних унаслідок диференціації клітин останніх.
Є кілька гіпотез походження еукаріотів, з яких у наш час найпопулярніша симбіотична, її послідовники вважають, що двомембранні органели, які мають свій геном і здатні до розмноження поділом (пластиди та мітохондрії) - нащадки симбіотичних прокаріотів, котрі втратили здатність до існування поза клітиною хазяїна. Спільне існування кількох видів прокаріотів привело врешті-решт до появи еукаріотичних клітин.
Ядро клітини.
Я
 дро
- найбільша
органела клітини, її найважливіший
регулюючий центр. Як правило, клітина
має одне ядро, але існують клітини
двохядерні і багатоядерні. У деяких
організмах можуть зустрічатися клітини,
позбавлені ядер. До таких без'ядерних
клітин відносяться, наприклад, еритроцити
ссавців,
тромбоцити, клітини ситовидних трубок
рослин і деякі інші типи клітин. Зазвичай
без'ядерними бувають високоспеціалізовані
клітини, що втратили ядра на ранніх
стадіях розвитку.
дро
- найбільша
органела клітини, її найважливіший
регулюючий центр. Як правило, клітина
має одне ядро, але існують клітини
двохядерні і багатоядерні. У деяких
організмах можуть зустрічатися клітини,
позбавлені ядер. До таких без'ядерних
клітин відносяться, наприклад, еритроцити
ссавців,
тромбоцити, клітини ситовидних трубок
рослин і деякі інші типи клітин. Зазвичай
без'ядерними бувають високоспеціалізовані
клітини, що втратили ядра на ранніх
стадіях розвитку.
Мал.1 Бузковим кольором забарвлені ядра клітин
Ядро забезпечує найважливіші метаболічні і генетичні функції клітини. Більшість клітин містять одне ядро, зрідка зустрічаються багатоядерні клітини (деякі гриби, прості, водорості, поперечно-смугасті м'язові волокна та ін.).
Ядро (лат. nucleus, грец. caryon — ядро) є вмістилищем генетичного матеріалу і місцем, де цей матеріал реалізується. Біологічне значення ядра визначається головним його компонентом — молекулами ДНК, здатними до реплікації та транскрипції. Ці дві властивості ДНК лежать в основі двох найважливіших функцій ядра клітини: (1) подвоєння спадкової інформації та передача її в ряді клітинних поколінь, (2) регулювання транскрипції (переписування інформації з ДНК на іРНК) і транспорт синтезованих всіх трьох видів РНК (іРНК, рРНК, тРНК) у цитоплазму клітини. Іншими словами, ядро є системою генетичної детермінації та регуляції білкового синтезу. Реплікація молекули ДНК дає можливість при мітозі двом дочірнім клітинам отримати якісно і кількісно однакові об’єми генетичної інформації.
Маса ДНК в інтерфазній клітині людини дорівнює ± 5 пг, в дрозофіли ±0,25 пг, але в клітинах тритона ± 95 пг. В ядрі є так звані репараційні ферменти, які ліквідують спонтанне пошкодження молекули ДНК. Клітина звичайно містить одне ядро, рідше 2–3 (у печінкових клітинах) або багато (в остеокластах).
Аналогом ядра в прокаріот є нуклеоїд (від лат. nucleus — ядро і грец. eides — подібний), сплетена “гола” (не оточена білками), циркулярна (замкнута) ДНК, занурена в цитоплазму. Нуклеоїд представлений ДНК, накрученою на стабілізаційні білки та РНК.
Розрізняють (1) інтерфазні ядра — звичайні ядра функціональних клітин, що не діляться, (2) метаболічні ядра і (3) ядра клітин під час мітозу.
Позбавлена ядра клітина швидко гине. Проте деякі клітини в зрілому (диференційованому) стані втрачають ядро. Такі клітини або живуть недовго і замінюються новими (наприклад, еритроцити), або підтримують свою життєдіяльність за рахунок припливу метаболитов з клітин, що тісно примикають до них, - "годувальник" (наприклад, клітини флоеми у рослин).
За формою ядро може бути кулястим, овальним, лопатевим, лінзовидним і так далі. Розмір, форма і структура ядер змінюються залежно від функціонального стану клітин, швидко реагуючи на зміну зовнішніх умов.
Ядро зазвичай переміщається по клітині пасивно із струмом цитоплазми, що оточує його, але іноді воно здатне самостійно пересуватися, здійснюючи рухи амебоїдного типу.
Форма ядра зазвичай округла або овальна. Діаметр ядра може коливатися від 5-10 мкм до 20 мкм. Зовні ядро оточене подвійною мембраною (ядерною оболонковою або кариолеммой), в якій є ядерні пори; через ці пори здійснюється зв'язок ядра з цитоплазмою.
Будова інтерфазного ядра.
Ядро - це найважливіша складова частина клітини, яка є практично у всіх клітинах багатоклітинних організмів. Більшість клітин має одне ядро, але бувають двоядерні та багатоядерні клітини (наприклад, поперечно-смугасті м'язові волокна). Двоядерних і багатоядерність обумовлені функціональними особливостями або патологічним станом клітин. Форма і розміри ядра дуже мінливі і залежать від виду організму, типу, віку і функціонального стану клітини. У середньому обсяг ядра складає приблизно 10% від загального об'єму клітини. Найчастіше ядро має округлу або овальну форму розміром від 3 до 10 мкм у діаметрі. Мінімальний розмір ядра становить 1 мкм (у деяких найпростіших), максимальний - 1 мм (яйцеклітини деяких риб і земноводних). У деяких випадках спостерігається залежність форми ядра від форми клітини. Ядро зазвичай займає центральне положення, але в диференційованих клітинах може бути зміщене до периферійного ділянці клітини. У ядрі зосереджена практично вся ДНК еукаріотичної клітини.
Основними функціями ядра є:
Зберігання та передача генетичної інформації;
Регуляція синтезу білка, обміну речовин і енергії в клітині.
Таким чином, ядро є не тільки вмістилищем генетичного матеріалу, але й місцем, де цей матеріал функціонує і відтворюється. Тому порушення будь-якої з цих функцій призведе до загибелі клітини. Все це вказує на провідне значення ядерних структур у процесах синтезу нуклеїнових кислот і білків.
Одним з перших учених продемонстрували роль ядра в життєдіяльності клітини був німецький біолог Хаммерлінг. В якості експериментального об'єкта Хаммерлінг використовував великі одноклітинні морські водорості Acetobularia mediterranea і А. c renulata. Ці близькоспоріднені види добре відрізняються один від одного за формою «шляпки». У підставі стеблинки знаходиться ядро. В одних експериментах капелюшок відокремлювали від нижньої частини стебла. В результаті було встановлено, що для нормального розвитку капелюшки необхідно ядро. В інших експериментах стеблинка з ядром одного виду водорості з'єднувався зі стеблинкою без ядра іншого виду. У утворилися химер завжди розвивалася капелюшок, типова для того виду, якому належало ядро.
Загальний план будови інтерфазних ядра однаковий у всіх клітин. Ядро складається з ядерної оболонки, хроматину, ядерець, ядерного білкового матриксу і каріоплазми (нуклеоплазми). Ці компоненти зустрічаються практично у всіх неделящіхся клітинах еукаріотичних одно-і багатоклітинних організмів.
а) будова та функції каріолеми, поняття про ядерні пори
Ядро клітини в інтерфазі, в період, коли вона не ділиться, характеризується наявністю таких складових, як каріолема (оболонка ядра), каріоплазма (ядерний сік), ядерця і хроматин, який є сукупністю інтерфазних хромосом тією чи іншою мірою деконденсованих, тобто потоншених.
Каріолема (від грец. karion — ядро i lema — оболонка), або нуклеолема (від лат. nucleus — ядро i lema — оболонка) чи поверхневий апарат ядра в інтерфазній клітині складається з трьох основних компонентів: ядерної оболонки, периферичної щільної пластинки і порових комплексів. Каріолема є спеціалізованою частиною загальної мембранної системи цитоплазми.
Найімовірніше, що каріолема утворилася з цистерн гранулярної ендоплазматичної сітки, містить лише 20–35% ліпідів і аж 65–80% білка. Каріолема утворена сплющеними цистернами і має відповідно зовнішню і внутрішню мембрани. Зовнішня мембрана безпосередньо переходить у мембрани грЕС і на своїй поверхні містить рибосоми (полісоми) та сітку проміжних (віментинових) філаментів. Між мембранами знаходиться перинуклеарний простір, завширшки від 10 до 100 нм, який з’єднується з порожнинами ендоплазматичної сітки. Під внутрішньою мембраною міститься ядерна пластинка (ламіна) — сітка проміжних філаментів, яка є периферичною частиною структурованого матрикса ядра. Вона супроводжує внутрішню мембрану ядерної оболонки і тісно зв’язана з білковими глобулами порового комплексу. Щільна ядерна пластинка (1) підтримує форму ядра, бере участь у: (2) впорядкованій укладці хроматину ядра, (3) структурній організації порових комплексів і (4) формуванні каріолем при поділі клітин.
б) каріоплазма
Каріоплазма (від грец. karion — ядро і plasma — виліплене) (рідше називають каріолімфою, або ядерним соком) — це білковий колоїд слабов’язкої консистенції. Мікроскопічно в ній виявляються грудочки хроматину (так звані хромоцентри). У каріоплазмі печінкових клітин щура знаходиться 92–98% білка глобулінової фракції, 12,4% гістонів, 22,2% ДНК і 2–8% РНК. Характер обміну в каріоплазмі досить своєрідний. Вважається, що в ядрах немає окисних ферментів (дегідрогеназ, цитохромоксидази, ферментів переносу електронів). У ній створюються умови для анаеробіозу, при яких хромосоми і функціонують. Крім цього каріоплазма містить глікоген.
Під світловим мікроскопом в ядрі видні хроматинові гранули, які є конденсованими ділянками хромосом. Конденсований хроматин недоступний для транскрипції отримав назву гетерохроматину (від грец. heteros — інший). Він міститься під каріолемою, навколо ядерця і розкиданий по каріоплазмі. Гетерохроматин поділяють на факультативний і конститутивний. Конститутивний, або структурний хроматин, ніколи не переходить в еухроматин, тоді як факультативний має здатність до такого перетворення. Останній включає ДНК, яка може піддаватися транскрипції, хоч в даний момент перебуває в конденсованому стані. При диференціації ядерних еритроцитів хребетних тварин різко збільшується кількість факультативного гетерохроматину.
Роль кожного з цих різновидів хроматину різна. Тоді як деконденсований хроматин, або еухроматин (від грец. eu — добрий) відкритий для транскрипції, бере участь у передачі генетичної інформації в інтерфазі, то конденсований хроматин, або гетерохроматин не виконує такої функції.
Функціональне значення і біохімічні характеристики каріоплазми, на жаль, ще не до кінця вивчені. Як і в матриксі цитоплазми, в каріоплазмі відбуваються реакції між глобулярними молекулами в суспензіях, тобто реакції найбільш численні і важливі, хоч досліджені неповністю.
в) ядерце
Ядерце виявляється в інтерфазному ядрі як гранула діаметром 1–2 мкм. У клітині звичайно буває 1-2, рідше більше ядерець, залежно від функціональної активності клітин, необхідності синтезу білка. Під електронним мікроскопом ядерце має вигляд електроннощільної губки з товстими шнурами, що анастомозують між собою, і гранулами речовини, інколи можуть бути видні вакуолі. Ядерце не має оболонки, контур його нерівний. За хімічним складом ядерце містить РНК і білки, зокрема багато кислих білків. В ядерці виявлені також фосфати та іони Ca, K, Mg, Fe, Zn.
Хроматин. Типи хроматину, його морфологічні та функціональні особливості
У каріоплазмі мікроскопічно спостерігається хроматин — речовина, яка добре забарвлюється основними фарбами. Хроматин — це матеріалхромосом, дуже довга спіралізована довголанцюгова нитка ДНК, зв’язана з деякою кількістю РНК, гістонів та інших основних білків. Загальна довжина молекул ДНК всіх хромосом в ядрі клітини людини становить понад 2 м, а в 5-періоді інтерфази 4 м. До цього часу неостаточно з’ясованими залишаються механізми, які оберігають ці нитки від переплутування. Проте очевидною є необхідність компактного упакування молекул ДНК, яке здійснюється гістоновими білками і забезпечує:
* упорядковане розміщення дуже довгих молекул ДНК в досить малому об’ємі ядра;
* функціональний контроль активності генів, який спрямовується завдяки особливостям упаковки окремих ділянок генів.
Під світловим мікроскопом в ядрі видні хроматинові гранули, які є конденсованими ділянками хромосом. Конденсований хроматин, недоступний для транскрипції, отримав назву гетерохроматину (від грецьк. heteros— інший). Він міститься під каріолемою, навколо ядерця і розкиданий по каріоплазмі. Гетерохроматин поділяють на факультативний і конститутивний. Конститутивний, або структурний, хроматин ніколи не переходить в еухроматин, тоді як факультативний має здатність до такого перетворення. Останній включає ДНК, яка може зазнавати транскрипції, хоч до цього перебувала в конденсованому стані. При диференціації ядерних еритроцилв хребетних тварин різко збільшується кількість факультативного гетерохроматину.
Роль кожного з цих різновидів хроматину різна. Якщо деконденсо- ваний хроматин, або еухроматин (від грецьк. eu — добрий), відкритий для транскрипції, бере участь у передачі генетичної інформації в інтерфазі, то конденсований хроматин, або гетерохроматин, не виконує такої функції.
Ядерце та його роль у клітині.
В ядрі є гліколітичні ферменти (гексокіназа і фосфофруктокіназа), які є АГФ-залежними, і такі, що приводять до утворення АТФ у другій стадії гліколізу (фосфогліцераткіназа і піруваткіназа). Другу групу ферменлв складають полімерази, а саме ДНК-полімераза, ДНК-лігаза (з’єднує фрагменти ДНК), ендонуклеази (вирізують ділянки І з,іпсова”ної ДНК), ДНК-залежна-РНК-полімераза (входить до складу ДНК і бере участь в синтезі високополімерної РНК).
Функціональне значення і біохімічні характеристики каріоплазми, на жаль, ще не до кінця вивчені. Як і в матриксі цитоплазми, в каріоплазмі відбуваються реакції між глобулярними молекулами в суспензіях, тобто реакції найбільш типові і важливі.
Ядерце виявляється в інтерфазному ядрі як гранула діаметром 1-2 мкм. У клітині зазвичай буває 1-2 (іноді більше) ядерець, залежно від функціональної активності клітин, необхідності синтезу білка. Під електронним мікроскопом ядерце має вигляд електронно-щільної губки з товстими шнурами, що анастомозують між собою, і гранулами речовини, інколи можуть бути видні вакуолі. Ядерце не має оболонки, контур його нерівний. За хімічним складом ядерце містить РНК і білки, зокрема, багато кислих білків. В ядерці виявлені також фосфати та іони Ca, K, Mg, Fe, Zn.
За допомогою електронного мікроскопа в ядерці виявлено: (1) гранулярну частину (15-20 нм в діаметрі), часто розташовану по периферії, — це попередники цитоплазматичних рибосом, (2) фібрили (завтовшки 5-8 нм), які є сукупністю первинних транскриптів рРНК, і (3) аморфну частину — скупчення петель ДНК, організаторів ядерець і специфічних РНК-зв’язаних білків. Фібрилярні та гранулярні компоненти ядерця формують так звану нуклеолонему — ядерцеву нитку завтовшки 60-80 нм, яка виділяється своєю щільністю на фоні менш щільного матриксу. Ядерце зникає в профазі і з’являється в телофазі: утворюється хромосомами — організаторами ядерець.
Функції ядерця — цитосинтез рРНК, а можливо, і тРНК, утворення або складання субодиниць рибосом, РНК ядерця можуть переходити в цитоплазму.
Структура та функція немембранних органел.
Д о
немембранних органел належать рибосоми,
мітохондрії, клітинний центр та органели
руху.
о
немембранних органел належать рибосоми,
мітохондрії, клітинний центр та органели
руху.
Що таке рибосоми? Які їхні функції? Рибосоми (від «рибонуклеїнова кислота» та грец. сома) - сферичні тільця діаметром приблизно 20 нм, які беруть участь у синтезі білків у клітині. Вони складаються з двох різних за розмірами субодиниць: великої та малої (мал. 5). Кожна з них містить рРНК і білки, які взаємодіють між собою.
Субодиниці рибосом можуть роз'єднуватись після завершення синтезу білкової молекули і знову сполучатись між собою у місцях синтезу білків.
Мал. 5. Схема будови рибосоми:
1 - мала субодиниця;
2 - велика субодиниця
Структурні компоненти рибосом утворюються в ядрі. Кількість рибосом у клітині залежить від інтенсивності процесів біосинтезу білків. Наприклад, у хребетних тварин найбільше рибосом виявлено в клітинах печінки, червоного кісткового мозку.
По локалізації підрозділяються на:
вільні (перебувають у гіалоплазмі);
невільні або прикріплені (пов'язані з мембранами ендоплазматичної мережі).
Кожна рибосома складається з малої й великий субодиниць. Кожна субодиниця рибосоми складається з рибосомальної РНК і білка рибонуклеопротеїа, які утворяться в ядерці. Складання субодиниць у єдину рибосому здійснюються в цитоплазмі. Для синтезу білка окремі рибосоми за допомогою матриці або інформаційної РНК поєднуються в ланцюжки рибосом - полісоми. Вільні й прикріплені рибосоми, крім відмінності в їхній локалізації, відрізняються певною функціональною специфічністю: вільні рибосоми синтезують білки для внутрішніх потреб клітини ( білки-ферменти, структурні білки), прикріплені синтезують білки "на експорт".
Цитоскелет. мікротрабекулярна сітка цитоплазми.
В цитоплазмі є внутрішній цитоскелет, що складається з мікротрабекулярної системи, мікротрубочок і мікрофіламентів. Мікротрабекулярна система являє собою мережу з тонких фібрил, які зв’язують всі внутрішньоклітинні компоненти.
Цитоплазма являє собою основну речовину клітини: цитоплазматичний матрикс (гіалоплазму) і мережу різних мембранних компонентів. Вона має вигляд своєрідних тривимірних решіток, побудованих з тонких білкових тяжів діаметром 3-6 нм, що заповнюють усю клітину. Усі компоненти цитоплазми виявляються немов підвішеними до цієї мікротрабекулярної решітки. Разом з мікротрубочками і мікрофіламентами вона складає так називнийцитоскелет клітини. Мікротрубочки – це тонкі органели діаметром 24 нм, побудовані з білка тубуліну, що сягають у довжину кількох мікрометрів. При розподілі клітини вони складають основу веретена поділу і також беруть участь у внутрішньоклітинному транспорті. Мікрофіламенти – це nt; тонкы былковы нитки (дыаметром 507 нм), що складаються з актино- і міозиноподібних білків. Вони утворюють рухову систему клітини при рухах цитоплазми чи деяких структур всередині неї.
Цитоскелет поділяє клітку на дві фази : багату білком і багату водою, що заповнює простір між тяжами. Обидві фази мають консистенцію гелю чи золю (більш рідкий стан), що може регулюватися самою клітиною. При цьому цікаво відзначити, що присутність іонів двохвалентних металів підвищує активність деяких білків, які у свою чергу підсилюють полімеризацію білків цитоскелету. У тому числі формування поперечних зшивок між ними. Це виявляється за збільшенням в’язкості цитоплазми. Подібна структура гіалоплазми пояснює ще одну її важливу властивість - явище тиксотропії. Тобто гіалоплазма знаходиться на грані між розчинністю і нерозчинністю.
Гіалоплазма не тільки підтримує внутрішню структуру клітини. Тут же протікають тисячі біохімічних реакцій гліколізу, біосинтезу і розпаду цукрі, ліпідів, нуклеїнових кислот, білків, тощо.
Рибосоми, структура та функції.
Рибосома (ribosome) є немембранною органелою клітини, що складається з рРНК тарибосомних білків (протеїнів). Рибосома здійснює біосинтез білків транслюючи мРНКполіпептидний ланцюг. Таким чином, рибосому можна вважати фабрикою, що виготовляє білки, базуючись на наявній генетичній інформації. В клітині дозрілі рибосоми знаходяться переважно в компартментах, для активного білкового синтезу. Вони можуть вільно плавати в цитоплазмі або бути прикріпленими до цитоплазматичного боку мембран ендоплазматичного ретикулуму чи ядра. Активні (ті що є в процесі трансляції) рибосоми знаходяться переважно у вигляді полісом. Існує ряд свідчень, які вказують на те, що рибосома є рибозимом.
Загальна будова
Рибосоми
прокаріотів
та еукаріотів
є дуже подібними за будовою та функцією,
але відрізняються розміром. Вони
складаються з двох субодиниць:
однієї великої та однієї малої. Для
процесу трансляції необхідна злагоджена
взаємодія обох субодиниць, що разом
становлять комплекс із молекулярною
масою декілька мільйонів дальтон
(Da). Субодиниці рибосом за звичай
позначаються одиницями Сведберга
(S), що є мірою швидкостіседиментації
під час центрифугування
і залежать від маси, розміру та форми
частинки. Позначені в цих одиницях,
велика субодиниця є 50S або 60S (прокаріотичні
або еукаріотичні, відповідно), мала є
30S або 40S, і ц
 іла
рибосома (комплекс малої разом з великою)
70S або 80S.
іла
рибосома (комплекс малої разом з великою)
70S або 80S.
Схематичне зображення рибосоми в процесітрансляції. Мала субодиниця зв'язує мРНК, а велика субодиниця напрямляє аміноацил-тРНК та пептидил-тРНК до матричної РНК, контролюючи правильне парування відповідних триплетів
Молекулярний склад
Молекулярний склад рибосом є доволі складним. Для прикладу, рибосома дріжджів ‘Saccharomyces cerevisiae’ складається із 79 рибосомних білків та 4 різних молекул рРНК.Біогенез рибосом є також надзвичайно складним і багатоступеневим процесом, що відбувається в ядрі та ядерці еукаріотичної клітини.
Атомна структура великої субодиниці (50S) організму Haloarcula marismortui була опублікована N. Ban et al. в журналі Science 11 Серпня 2000 року. Невдовзі після цього, 21шого Вересня 2000 року, B.T. Wimberly, et al., опублікували в журналі Nature структуру 30S субодиниці організму Thermus thermophilus. Використовуючи ці координати, M.M. Yusupov, et al. зуміли реконструювати цілу 70S частинку Thermus thermophilus і опублікувати її в журналі Science, в Травні 2001. У 2009 році професор Джордж Чьорч (George Church) та коллеги з Гарвардустворили повністю функціональну штучну рибосому в звичайних умовах, які присутні в клітинному середовищі. Як конструкційні елементи використовувались молекули з розщепленої за допомогою ензимів кишкової палички. Створена рибосома успішно синтезує білок, що відповідає за біолюмінесценцію.
Функція
Рибосома є органелою, на якій відбувається трансляція генетичної інформації закодованої в мРНК. Ця інформація втілюється в синтезований тут-жеполіпептидний ланцюг. Рибосома несе двояку функцію: є структурною платформою для процесу декодування генетичної інформації з РНК, та володієкаталітичним центром відповідальним за формування пептидного зв’язку, так званим ‘пептидил-трансферазним центром’. Вважається що пептидил-трансферазна активність асоціюється з рРНК, і тому рибосома є рибозимом.
Включення та їх типи.
Включення — це непостійні утвори в цитоплазмі, представлені у вигляді гранул, крапельок чи кристаликів, які служать для забезпечення життєдіяльності клітини, або з’являються в результаті її функціонування. Немає стрункої класифікації включень, тому, залежно від природи речовини, яка в них нагромаджується, і для зручності вивчення їх традиційно поділяють на такі групи:
1) трофічні: білкові включення частіше містяться в рослинах, наприклад, алейронові зерна в клітинах насіння рослин. Із вуглеводних трофічних включень найбільше розповсюдженими є гранули полісахариду глікогену в печінці. Ліпідні включення мають вигляд крапель, які можуть зливатися в жирових клітинах сполучної тканини;
2) пігментні включення — це речовини, які мають певне забарвлення. Сюди відносять гемоглобін в еритроцитах, гемосидерин — продукт обміну гемоглобіну, який нагромаджується в макрофагах, меланін — чорний пігмент — синтезується і нагромаджується в пігментоцитах шкіри, ліпофусцин (“пігмент старіння”) збирається у вигляді мембранних гранул у нервових клітинах;
3) секреторні включення — міхурці, в яких збираються ферменти, продукти синтезу залозистих клітин, наприклад, крапельки слини в клітинах слинних залоз, гранули зимогену в клітинах підшлункової залози;
4) екскреторні включення містять непотрібні, а навіть шкідливі продукти метаболізму, які підлягають видаленню з клітини (і з організму) наприклад, сечова кислота (у клітинах нирки), жовчні пігменти (у печінкових клітинах);
5) специфічні — група включень, які виконують особливу роль в клітинах: вітамін С в сітчастому шарі кори надниркових залоз, кератогіалін — білок у зернистому шарі епітелію шкіри, елеідин — білок в блискучому шарі епітелію шкіри тощо;
6) неспецифічні — невластиві живій нормальній клітині (частинки туші при татуюванні, включення вугілля, оксиду заліза, кремнію — у клітинах легень).
Високоспеціалізовані компоненти цитоплазми диференційованих клітин.
Ян Пуркіньє встановив, що живим компонентом клітини є внутрішній вміст, який він назвав «протоплазмою». Для встановлення функцій окремих компонентів клітини важливо виділити їх у чистому вигляді, найчастіше це робиться за допомогою методу диференційного центрифугування. Отримання фракцій клітинних органел починається із руйнування плазмалеми. Утворений гомогенат послідовно центрифугується при різних швидкостях. На першому етапі можна отримати чотири фракції: (1) ядер і великих уламків клітин, (2) мітохондрій, пластид, лізосом і пероксисом, (3) міркосом — пухирців апарату Гольджі та ендоплазматичного ретикулуму, (4) рибосом. У супернатанті залишаться білки та дрібніші молекули. Подальше диференційне центрифугування кожної із змішаних фракцій дозволяє отримати чисті препарати органел, до яких можна застосовувати різноманітні біохімічні та мікроскопічні методи.
Багатоклітинні організми складаються із клітин, що тією чи іншою мірою відрізняються за будовою і функціями, наприклад у дорослої людини близько 230 різних типів клітин[55]. Всі вони є нащадками однієї — зиготи (у випадку статевого розмноження) — і набувають відмінностей внаслідок процесу диференціації. Диференціація у переважній більшості випадків не супроводжується зміною спадкової інформації клітини, а забезпечується лише шляхом регулювання активності генів, специфічний характер експресії генівуспадковується під час поділу материнської клітини зазвичай завдяки епігенетичниммеханізмам. Проте є винятки: наприклад при утворенні клітин специфічної імунної системихребетних відбувається перебудовування деяких генів, еритроцити ссавців повністю втрачають всю спадкову інформацію, а статеві клітини — її половину.
Відмінності між клітинами на перших етапах ембріонального розвитку з'являються по-перше внаслідок неоднорідності цитоплазми заплідненої яйцеклітини, через яку під час процесу дроблення утворюються клітини, що різняться за вмістом певних білків та РНК, по-друге, важливу роль відіграє мікрооточення клітини — її контакти з іншими клітинами та середовищем.
Диференціюючись, клітини втрачають свої потенції, тобто здатність давати початок клітинам інших типів. Із тотипотентих клітин, до яких належить зокрема зигота, може утворитись цілісний організм. Плюрипотентні клітини (наприклад клітини бластоцисти) мають можливість диференціюватись у будь-який тип клітин організму, але з них не можуть розвинутись позазародкові тканини, а отже і нова особина. Клітини, які здатні дати початок тільки обмеженій кількості інших тканин називаються мультипотентними (стовбурові клітинидорослої людини), а ті, які можуть відтворювати тільки собі подібні — уніпотентними. Багато із остаточно диференційованих клітин (наприклад нейрони, еритроцити) повністю втрачають здатність до поділу і виходять з клітинного циклу.
У деяких випадках диференціація може бути зворотною, протилежний до неї процес називається дедиференціація. Він характерний для процесів регенерації, але інколи може відбуватись патологічно, як етап злоякісної трансформації клітин
Диференціація клітин відбувається, починаючи з ранніх стадій ембріогенезу, і продовжується формуванням тканин. Під час диференціації наступають зміни в цитоплазмі клітин внаслідок її взаємодії з ядром. Проте, найбільш помітною є морфологічна диференціація.
У ході розвитку виділяють ряд етапів (типів) морфологічної диференціації:
1) Оотипічна, яка проявляється вже під час запліднення. Особливо наглядно вона виступає в зиготі амфібій. На анімальному полюсі появляється так званий сірий серп, а в базальному — нагромаджується жовток.
2) Бластомерна — на рівні перших декількох дроблень. У деяких тварин чітко помітна різниця між окремими групами бластомерів, у жаби — це мікробластомери і макробластомери.
3) Зачаткова. У період утворення мезодерми відзначається різниця між клітинами окремих зачатків, коли в сомітах вони інші, ніж у сегментних ніжках чи спланхнотомі.
4) Тканинний тип диференціації продовжується весь період ембріонального і навіть постнатального розвитку. Найбільш помітна така диференціація в процесі розвитку мезенхіми, з якої утворюються всі види сполучної тканини (пухка, щільна, хрящова, кісткова) і формені елементи крові (еритроцити, всі види лейкоцитів).
Детермінація. У ході диференціації клітини певної тканини реалізують закріплені детермінацією потенції. Детермінація клітин(тканин) (від лат. determinatio — визначення) — процес, який програмує напрямок розвитку клітин, характерний для даного виду тканин. У результаті детермінації компетентна клітинна система обирає один з багатьох можливих шляхів розвитку. Отже, детермінація клітини — це процес визначення шляху її дальшого розвитку, виникнення якісної своєрідності клітин.
Клітина, детермінована в одному напрямку, продукує клітини, детерміновані таким же чином: “подібне породжує подібне”. Немає єдиної теорії механізму детермінації. Вважають, що детермінація відбувається внаслідок дії низки факторів. Оскільки в процесі детермінації не має значення блокування генів, бо під час реплікації ДНК всі гени повинні бути деблоковані, то виникає думка, що детермінованість у різних клітинних лініях мабуть залежить від постійності цитоплазми протягом послідовних поколінь. Цитоплазма впливає певним чином на активацію або гальмування генів, що приводить до синтезу певних молекул РНК, а тим самим і синтезу білків відповідних даному типові клітин.
Розмноження соматичних клітин.
Соматичні клітини складають популяцію, утворену при безстатевому розмноженні (поділ клітин). Соматичні мутації обумовлюють генотипну різноманітність тканин, часто не передаються по спадковості і обмежені тим індивідуумом, у якому вони виникли. Соматичні мутації виникають у диплоїдних клітинах, тому проявляються тільки за домінантними генами або за рецесивними, але в гомозиготному стані. Чим раніше в ембріогенезі людини виникла мутація, тим більша ділянка соматичних клітин відхиляється від норми. І навпаки, чим пізніше в процесі розвитку організм зазнає мутаційного впливу, тим менша та ділянка тканини, яка утворюється із мутаційної клітини. Наприклад, забарвлення райдужної оболонки ока - білий або карий сегменти на блакитній райдужці — зумовлені соматичною мутацією. Вважають, що наслідком соматичних мутацій є ракове переродження. Злоякісний ріст викликається канцерогенами, серед яких найбільш негативні - проникаюча радіація й активні хімічні сполуки (речовини).І хоча соматичні мутації не успадковуються, вони знижують репродуктивні можливості організму, в якому виникли.Мутації, що виникають в гаметах або в кліти¬нах, з яких вони утворюються, отримали назву генеративних, або термінальних мутацій. Чим раніше в статевих клітинах виникає мутація, тим більшою буде частка статевих клітин, які нестимуть нову мутацію. Верхня межа частки клітин, які будуть містити індуковану або спонтанну мутацію, складає 50 відсотків. Існує думка, що найбільша кількість мутацій у статевих клітинах виникає в овоцитах. Оскільки сперматогонії зазнають постійного поділу, то серед них може відбуватися відбір проти мутацій, які зумовлюють шкідливий ефект, і частота мутацій знижується до часу статевої зрілості. Жінка, навпаки, народжується майже з усіма мутантними змінами, в лінії статевих клітин немає паралельного мітотичного відбору. Овоцити не тільки не зазнають мітозу, вони залишаються малоактивними впродовж десятиліть, доки не стануть яйцеклітинами. За цей період овоцити старіють, стають непропорційно чут¬ливими до спонтанної мутації.На статеві клітини найбільший вплив спричиняють цезій-137, стронцій-90 і вуглець-14.Генеративні мутації при статевому розмноженні передаються наступним поколінням. Домінантні мутації з'являються вже в першому поколінні, а рецесивні - тільки в другому і наступних поколіннях, з переходом у гомозиготний стан.
Соматичні мутації зумовлюють генотипну різноманітність тканин однієї особини і здебільшого не передаються по спадковості при статевому розмноженні. При безстатевому розмноженні, якщо організм розвивається з однієї клітини або групи клітин, в яких виникла мутація, - такі зміни можуть передаватися нащадкам. Соматичні мутації вико¬ристовують у організмів, які розмножуються вегетативно. Соматичні мутації складають основу селекції культурних рослин, зокрема цитрусових.
Відомі дві основні форми розмноження рослин і тварин: статева і безстатева. Між ними існує принципова відмінність, яка полягає в тому, що при безстатевому розмноженні нове покоління бере початок лише від однієї батьківської особини, причому джерелом утворення нового покоління є соматичні (вегетативні) клітини.
У разі статевого розмноження новий організм, як правило, утворюється від двох батьківських особин (чоловічої і жіночої). Джерелом утворення цього організму є особливі клітини, які називають статевими, або гаметами. Від соматичних клітин вони відрізняються половинним (гаплоїдним) набором хромосом.
У разі вегетативного розмноження новий організм бере початок не із спеціалізованих, а із звичайних соматичних клітин. Цей тип розмноження особливо поширений у рослин.
Клітинний (мітотичний) цикл.
К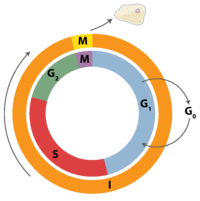 літинний
цикл — це серія подій, що відбувається
у період від утворення еукаріотичної
клітини до завершення її поділу.
Клітинні поділи необхідні як утворення
тіла багатоклітинних організмів,
так і для відтворення собі подібних.
Перед поділом генетичний
матеріал має бути репліковано,
щоб кожна з нових клітин отримала його
копію, ідентичну до материнської.
літинний
цикл — це серія подій, що відбувається
у період від утворення еукаріотичної
клітини до завершення її поділу.
Клітинні поділи необхідні як утворення
тіла багатоклітинних організмів,
так і для відтворення собі подібних.
Перед поділом генетичний
матеріал має бути репліковано,
щоб кожна з нових клітин отримала його
копію, ідентичну до материнської.
Середня тривалість клітинного циклу еукаріотичної клітини за сприятливих умов і наявності стимулів до поділу може становити 24 год. Він складається із наступних фаз:
Інтерфаза — період, коли клітина не ділиться; триває 90% часу клітинного циклу і в свою чергу поділяться на три фази:
Фаза G1 (англ. first gap) — пресинтетичний період, клітина росте, накопичує поживні речовини, виконує свої основні функції (5—6 год або більше залежно від типу клітин та умов).
Фаза S — синтетичний період, відбувається реплікація ДНК, продовжується ріст клітини (10—12 год для людської клітини).
Фаза G2 (англ. second gap) — постсинтетичний період, клітина готується до поділу — перевіряє, чи добре скопійовано ДНК, накопичує білки необхідні для утворення веретена поділу, подвоюються деякі органели (4—6 год для типової людської клітини).
Клітинний поділ — триває не більше години і поділяється на два взаємопов'язані етапи:
Мітоз — поділ ядра, під час якого відбувається рівномірний розподіл генетичної інформації. Відбувається у кілька етапів: профаза, прометафаза, метафаза, анафазаі телофаза. Під час мітозу спіралозвані хромосоми, що складаються із двох ідентичних хроматид вишиковуються по екватору веретена поділу, а потім окремі хроматиди, за допомогою мікротрубочок, розходяться до його полюсів. На кожному полюсі формується нове ядро.
Цитокінез — поділ цитоплазми клітини, у тварин відбувається за участі скоротливого кільця із актинових та міозинових філамтенів, а у вищих рослин — за допомогою спеціальної структури — фрагмопласту, що складається із мікротрубочок веретена поділу і везикул апарату Гольджі, які, зливаючись між собою, відокремлюють дві дочірні клітини[51].
Окрім мітозу існує ще один спосіб поділу ядра еукаріотичної клітини — мейоз, це серія із двох поділів, між якими часто відсутня інтерфаза. На противагу мітозу, після завершенню мейозу кожна дочірня клітина отримує лише половину генетичної інформації батьківської клітини. Мейоз обов'язково відбувається на певному етапі життєвого циклу всіх організмів, здатних до статевого розмноження. Він необхідний для підтримання сталої кількостіхромосом у всіх особин виду і для здійснення генетичної рекомбінації — перегрупування та перерозподілу генів.
У багатоклітинних організмів частина диференційованих клітин виходять із клітинного циклу: після стадії G1 вони переходять у стадію спокою — G0, більшість таких клітин за певних умов можуть відновлювати проліферацію.
Усі події у клітинному циклі чітко регулюються системою спеціальних білків циклінів та циклін-залежних кіназ, яка тісно пов'язана з іншими сигнальними шляхами клітини. Якщо один або кілька елементів цієї системи виходять із ладу, це може призвести до неконтрольованого поділу клітин і утворення пухлин, зокрема, злоякісних[48].
Інтерфаза та її періоди.
І нтерфаза
(від лат. інтер - між і грец. фазіс - поява)
- період між двома послідовними поділами
клітини або від завершення останнього
поділу до її загибелі (наприклад, клітини
багатоклітинних організмів, які втрачають
здатність до поділу).
нтерфаза
(від лат. інтер - між і грец. фазіс - поява)
- період між двома послідовними поділами
клітини або від завершення останнього
поділу до її загибелі (наприклад, клітини
багатоклітинних організмів, які втрачають
здатність до поділу).
В інтерфазі клітина росте, в ній подвоюються молекули ДНК, мітохондрії, пластиди, синтезуються білки й інші органічні сполуки. В цей період активно запасається енергія, потрібна для наступного поділу клітини.
Процеси синтезу найінтенсивніше відбуваються в певний період інтерфази, який називають синтетичним (мал. 69). У цей час подвоюються хроматиди (процес, пов'язаний з подвоєнням молекул ядерної ДНК). Проміжок часу між завершенням попереднього поділу клітини і синтетичним періодом називають
передсинтетичним, а між завершенням синтетичного періоду і початком наступного поділу - постсинтетичним (мал. 69).
Тривалість інтерфази, звичайно, становить до 90% часу всього клітинного циклу. Стимулом для наступного поділу клітини є досягнення нею певних розмірів у інтерфазі.
Основним способом поділу еукаріотичних клітин є мітоз (мал. 70). Процес мітозу (від грец. мітос — нитка) супроводжується ущільненням хромосом і утворенням особливого апарату, який забезпечує рівномірний розподіл спадкового матеріалу материнської клітини між двома дочірніми.
Які фази розрізняють під час мітозу? Мітоз складається з чотирьох послідовних фаз: профази, метафази, анафази і телофази. Він триває від кількох хвилин до 2-3 годин (мал. 70).
Мал. 70. Схема мітозу: профаза (1); метафаза (2); анафаза (3); телофаза (4)
Профаза (від грец. про - перед, раніше та фазіс) починається з ущільнення хроматину. Внаслідок цього під світловим мікроскопом можна розглянути будову хромосом і підрахувати їхню кількість. При цьому хроматиди вкорочуються і потовщуються, тобто спіралізуються. Стає помітною і первинна перетяжка, де розташована центромера, до якої приєднуються нитки веретена поділу. Поступово зменшуються в розмірах і зникають ядерця, ядерна оболонка розпадається на фрагменти, і хромосоми опиняються в цитоплазмі.
У цей час починає формуватися веретено поділу. Його нитки прикріплюються до центромер і хромосоми починають рухатись до центральної частини клітини.
Під час наступної фази мітозу - метафази (від грец. мета - після, через) - завершується спіралізація хромосом і формування веретена поділу. Хромосоми «вишиковуються» в одній площині в центральній частині клітини. При цьому їхні центромери розташовані на однакових відстанях від полюсів клітини. Наприкінці метафази хроматиди кожної хромосоми відокремлюються одна від одної.
Анафаза (від грец. ана - знову, поза) - найкоротша фаза мітозу. В цей час діляться центромери і хроматиди розходяться до різних полюсів клітини. Кожна з хроматид відповідає половині профазної хромосоми, тобто вони містять ідентичний спадковий матеріал.
Телофаза (від грец. телос - кінець) триває від моменту припинення руху хроматид до утворення двох дочірніх клітин. На початку телофази хромосоми
деспіралізуються. Навколо кожного з двох скупчень хроматид формується ядерна оболонка, з'являються ядерця і ядра дочірніх клітин набувають вигляду інтер-фазних. Протягом цієї фази поступово зникає веретено поділу. Наприкінці телофази ділиться цитоплазма материнської клітини і утворюються дві дочірні.
Цей процес відрізняється в клітинах рослин і тварин. У цитоплазмі рослинних клітин між дочірніми ядрами формуються клітинні стінки (як ви пригадуєте, в цьому процесі бере участь комплекс Гольджі). В клітинах тварин плазматична мембрана впинається всередину цитоплазми, утворюючи перетяжку, яка ділить клітину навпіл.
Мітоз та його стадії. Амітоз.
Мітоз (рідше: каріокінез або непрямий поділ) (від грец. mitos — нитка) — поділ ядра [етапкон'югацію гомологічних хромосом протягом профази.
Мітоз — лише одна з частин клітинного циклу, але він достатньо складний, щоб в його складі, у свою чергу, було виділено чотири фази: профаза, метафаза, анафаза і телофаза. Подвоєння хромосом відбувається ще в ході інтерфази. В результаті цього, в мітоз хромосоми вступають вже подвоєними, такими, що нагадують букву «X» (ідентичні копії материнської хромосоми сполучені один з одним в області центромери).
У профазі відбувається конденсація хромосом, клітинний центр ділиться і продукти його поділу розходяться до полюсів ядра, руйнується ядерна оболонка, утворюється веретено поділу.
У метафазі хромосоми розташовуються на «екваторі» (на рівній відстані від «полюсів» ядра) в одній площині, утворюючи так звану метафазну пластинку. Важливо відзначити, що вони залишаються в такому положенні протягом досить тривалого часу. Зазвичай у зв'язку з цим метафаза — найслушніший час для підрахунку хромосомних чисел.
У анафазі, яка є найкоротшою фазою мітозу, хромосоми діляться (з'єднання в районі центромери руйнується) і розходяться до різних полюсів клітини.
У телофазі відбувається руйнування веретена поділу і утворення ядерної оболонки навколо дочірніх ядер.
Тривалість мітозу в середньому 1-2 год., різна для різних видів клітин. Процес залежить також і від умов зовнішнього середовища (температури, світлового режиму й інших показників).
АМІТÓЗ (грец. а- — заперечення + mitos — нитка) — поділ соматичних клітин, який відбувається без спіралізації хромосом і без утворення веретена поділу (відрізняється від мітозу, де відбувається прямий поділ інтерфазного ядра клітини, а згодом іцитоплазми). Під час амітозу ядерце видовжується, перешнуровується, а потім витягується і ядро. У деяких випадках в ядрі виникає перегородка, що ділить його на дві частини. Поділ ядра іноді супроводжується поділом цитоплазми.
Розрізняють кілька форм амітозу:
рівномірний, коли утворюється два рівних ядра;
нерівномірний, коли утворюються нерівні ядра;
фрагментаційний, коли ядро розпадається на багато дрібних ядер однакової або різної величини.
Чи відбувається попередній синтез ДНК перед початком амітозу і як вона розподіляється між дочірніми ядрами — невідомо. Іноді при поділі певних клітин мітоз чергується з амітозом.
Амітоз — це своєрідний тип поділу, що іноді спостерігається при нормальній життєдіяльності клітини, а здебільшого при порушеннях функції, часто під впливом опромінення чи дії інших шкідливих чинників. Він властивий високодиференційованим клітинам. Амітоз, порівняно з мітозом, зустрічається рідше і відіграє другорядну роль у клітинному поділі переважної більшості живих організмів.
При амітозі ядро клітини без попередніх структурних змін ділиться на дві або більше частин. При цьому, на відміну від непрямого поділу клітини — каріокінезу, або мітозу, хромосоми з ядерної речовини не утворюються. Після поділу ядра звичайно ділиться цитоплазма.
Диференціація, старіння і смерть клітини.
У процесі життєдіяльності, після досягнення певного віку клітини старіють. Механізми клітинного старіння залишаються нез’ясованими. Згідно з однією гіпотезою, старіння є результатом катастрофічного нагромадження помилок біосинтетичних механізмів клітини, згідно з іншою — воно є наслідком обмеження можливостей росту клітин. Вважають, що старіння клітин є механізмом стабілізації кількості клітин у дорослому організмі.
Тривалість життя клітин у різних тканинах дорослого організму людини неоднакова. Клітини деяких тканин живуть дуже коротко — від декількох хвилин (лейкоцити) до декількох діб (клітини кишкового епітелію), чи багатьох років (кардіоміоцити і нейрони). У фізіологічних умовах старіння клітин закінчується програмованою смертю — апоптозом.
Клітини, як і все живе, гинуть. Смерть клітин може наступати як у нормі, так і при патології. Гинуть клітини вже під час ембріонального розвитку при формуванні тканин і органів. У ранньому онтогенезі наступає масова загибель певних груп клітин, які не досягли стану остаточної (термінальної) диференціації, наприклад, нейробласти. У дорослому організмі смерть клітин наступає: (1) внаслідок старіння, (2) при втраті функцій і (3) під впливом шкідливих факторів (некроз). Гинуть клітини, які виконали свою функцію. Так, смерть еозинофілів наступає після їх дегрануляції.
Некроз. Апоптоз.
Апоптоз (від грец. apoptosis — листопад, опадання) — природна (запрограмована, фізіологічна) смерть клітин; це активний, генетично контрольований процес загибелі клітин, який регулюється внутрішньоклітинною програмою, а запускається зовнішніми факторами. Апоптоз називається “смертю клітини в результаті самознищення”. Особливістю апоптозу є те, що він наступає в окремих клітинах або в їх групах, розділених значними угрупуваннями життєздатних клітин. Апоптоз спостерігається в різних клітинах людини і тварин в нормі, при патології, як у дорослих особин, так і під час ембріонального розвитку.
Зміни клітин при апоптозі. На початковій (латентній) стадії в клітині відзначається синтез ферментів, необхідних для забезпечення її загибелі. Найбільш раннім проявом апоптозу є втрата міжклітинних з’єднань. Після цього під впливом активованої ендонуклеази наступає мікрофрагментація хроматину і його маргінізація (переміщення під каріолему у вигляді півмісяців). Відтак наступає фрагментація ядра. Цитоплазма конденсується, мембранні органели спочатку розширяються, а відтак розпадаються з утворенням кластерів. На поверхні клітин утворюються здуття. Під кінець апоптозу за участю актинових мікрофіламентів відшнуровуються ділянки цитоплазми і фрагменти ядра, і таким чином утворюються апоптозні тіла різних розмірів, які руйнуються остаточно: одні можуть поглинатися сусідніми клітинами, інші — захоплюються фагоцитами. Реакції запалення при апоптозі не виникає. Нейтрофіли в фагоцитозі апоптозних тіл участі не беруть.
Апоптозний процес триває досить коротко — від декількох хвилин до декількох годин (у середньому від 1 до 3 годин). Латентний період, який передує самому процесові апоптозу, продовжується значно довше — до 12 годин. Але не всі клітини, в яких почався латентний період, вступають в апоптоз.
Механізми запуску і розгортання процесів апоптозу. Програмована смерть клітин можливо індукується нагромадженням генетичних помилок або знищенням чутливості клітин до ростових сигналів, а ініціюється рядом сигналів і ферментів. На початковому етапі (латентному періоді) синтезуються ферменти, необхідні для здійснення загибелі клітин. Але не всі клітини з активованими ферментами гинуть, частина з них виживає завдяки активації генів-рятувальників і трофічних факторів. Дальше процес апоптозу розгортається під впливом сигналів, які передаються до ядра (фактори, опосередковані іонами Са2+, ферменти), активацієюлетальних, або “кілерних” генів і шляхом синтезу апоптоз-специфічних білків. Важливу роль в розгортанні апоптозу мають ферменти: протеази та ендонуклеази, які зумовлюють мікрофрагментацію ДНК у ділянках між нуклеосомами.
Значення апоптозу. Апоптоз є одним із функціональних біологічних механізмів розвитку тканин і тканинного гомеостазу. Він до певної міри пов’язаний з усіма проявами життєдіяльності клітин у нормі та патології.
Особливе значення апоптозу в ембріональному розвиткові, який полягає в забезпеченні регресії частин ембріональних зачатків, особливо під час гісто- і органогенезу. Ембріональний розвиток завжди супроводжується надмірним утворенням клітин, особливо попередників нейронів (нейробластів), а під впливом апоптозу гине від 25 до 85% цих клітин. Проявами апоптозу в ембріогенезі є зміна форми закладок органів, розрив плодових оболонок.
У дорослому організмі в фізіологічних умовах старіння клітин завершується програмованою смертю. (а) У процесі імунної відповіді гинуть клітини-кілери. Клітини підгрудинної залози, які виконували гормональну функцію в дитячому віці, при настанні статевої зрілості також гинуть. Апоптоз клітин наступає при інволюції гормонально залежних органів після припинення гормональної стимуляції (постлактаційна інволюція молочної залози). (б) У клітинах імунної системи апоптоз забезпечує розвиток імунних реакцій (інволюція підгрудинної залози при настанні статевої зрілості). При дії на організм шкідливих факторів, при інфаркті, інсульті та при інфекційних захворюваннях наступає також апоптоз.
Слід відзначити, що апоптоз у зрілому організмі забезпечує клітинний гомеостаз — відносну постійність складу тканин і органів, а при старінні чи патології здійснює регуляцію кількості клітин в органах відповідно до знижених функціональних можливостей організму. Вважають, що пригнічення апоптозу можливо служить одним із механізмів канцерогенезу — злоякісного росту. Таке передбачення базується на тому, що в пухлинних клітинах інактивуються фактори, які запускають і підтримують програму апоптозу.
в) некроз
Некроз (від грец. nekrosis — змертвіння, вмирання) — загибель у результаті незворотного пошкодження клітин або ділянки тканини, органу. Загибель клітин наступає внаслідок дії різко виражених шкідливих факторів: перегріванні, переохолодженні, нестачі кисню (гіпоксії), порушенні кровопостачання (ішемії), дії отрут, хімічних препаратів, механічної травми тощо. При некрозі найчастіше помітні руйнування ядра: пікнози, каріолізиси і каріорексиси.
На початкових етапах розвитку некрозу наступає набряк цитоплазми та окремих органел (особливо мітохондрій), дисперсія рибосом і розширення цистерн ЕС. При цьому збільшується кількість лізосом, нагромаджуються жирові і пігментні включення, наростає проникність клітинних мембран, вакуолізація цитоплазми і ядра. У гіалоплазмі зростає концентрація Са2+, що приводить до активації фосфорилаз і руйнування мембранних фосфоліпідів та пошкодження мембран. На пізніх стадіях некрозу з лізосом виділяються ферменти і руйнують структури клітини.
При тому під впливом лізосомальної ДНКази фрагментується ДНК і гетерохроматин конденсується під каріолемою у вигляді великих грудок. Відтак ядро зменшується, ущільнюється і піддається каріопікнозу (від грец. karyon — ядро і pyknosis — ущільнення), потім гетерохроматин розпадається на частини, і таке явище називається каріорексисом (від грец. karyon — ядро і rhexis — розрив). Завершуються зміни в ядрі каріолізисом (від грец. karyon — ядро і lysis — розчинення) — остаточним його руйнуванням, ніби його розчиненням. У подальшому розриваються мембрани клітини і вона розпадається. Продукти розпаду клітини потрапляють у міжклітинний простір і фагоцитуються лейкоцитами і макрофагами. Остаточне руйнування клітини супроводжується запальним процесом, викликаним продуктами розпаду клітини.
Статева система і розвиток статевих клітин.
Людина підлягає усім біологічним законам розмноження вищих ссавців. Але функція розмноження людини підлягає ще соціальним законам, які в цивілізованому суспільстві не суперечать біології людини, а сприяють формуванню здорового, гармонійно розвинутого потомства.
Здібність до самовідтворення — неодмінна умова збереження видів. Тривалість життя кожної особини обмежена. У результаті розмноження одне покоління особин зміняє інше, забезпечуючи продовження існування виду. В процесі еволюції основним способом розмноження стає статевий, який можливий завдяки розвитку в обох статей спеціальної системи.
У ядрі кожної клітини є структури, які забезпечують під час її поділу пере дачу спадкової інформації дочірнім клітинам, а також реалізацію цієї інформації в процесі індивідуального розвитку. Це хромосоми (від грец. хрома - колір і сома - тіло). У ядрах клітин тіла людини міститься 46 хромосом, в яких закладена повна спадкова інформація. Такий набір хромосом називають диплоїдним і позначають 2n. Статеві клітини (гамети) несуть половину спадкової інформації, тому в них по 23 хромосоми. Це гаплоїдний набір хромосом - 1n. Диплоїдний набір хромосом відновлюється під час запліднення (злиття чоловічої і жіночої статевих клітин) й утворення зиготи.
Я
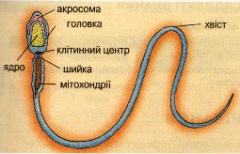 йцеклітини
та сперматозоїди розвиваються з первинних
статевих
клітин. Чоловічі первинні статеві
клітини починають утворюватися з
настанням статевої зрілості. Процес
дозрівання сперматозоїдів триває понад
два місяці. Щодня у чоловіка дозріває
кілька мільйонів сперматозоїдів. На
відміну від чоловіків, у жінок первинні
статеві клітини утворюються вже в
ембріональному періоді й перебувають
у «законсервованій» стадії аж до настання
статевої зрілості.
йцеклітини
та сперматозоїди розвиваються з первинних
статевих
клітин. Чоловічі первинні статеві
клітини починають утворюватися з
настанням статевої зрілості. Процес
дозрівання сперматозоїдів триває понад
два місяці. Щодня у чоловіка дозріває
кілька мільйонів сперматозоїдів. На
відміну від чоловіків, у жінок первинні
статеві клітини утворюються вже в
ембріональному періоді й перебувають
у «законсервованій» стадії аж до настання
статевої зрілості.
Ч
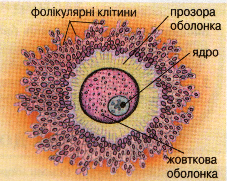 оловічі
статеві клітини - сперматозоїди - це
дуже дрібні, видовжені, рухливі гамети,
що складаються з головки, шийки і хвоста
(мал.1). Головка має ядро з гаплоїдним
набором хромосом, оточене тонким
шаромцитоплазми,
і апікальне тільце (акросому), яка
виробляє фермент, що сприяє проникненню
сперматозоїда в яйцеклітину. У шийці
розміщені клітинний центр (центріолі)
та велика кількість мітохондрій, енергія
яких забезпечує рух хвоста, а отже, і
рух самого сперматозоїда до яйцеклітини.
оловічі
статеві клітини - сперматозоїди - це
дуже дрібні, видовжені, рухливі гамети,
що складаються з головки, шийки і хвоста
(мал.1). Головка має ядро з гаплоїдним
набором хромосом, оточене тонким
шаромцитоплазми,
і апікальне тільце (акросому), яка
виробляє фермент, що сприяє проникненню
сперматозоїда в яйцеклітину. У шийці
розміщені клітинний центр (центріолі)
та велика кількість мітохондрій, енергія
яких забезпечує рух хвоста, а отже, і
рух самого сперматозоїда до яйцеклітини.
Мал.1. Будова сперматозоїда
Жіночі статеві клітини - яйцеклітини, на відміну від сперматозоїдів, нерухомі, значно більші за розміром, - кулястої форми (мал.2). Цитоплазма яйцеклітини містить великий запас поживних речовин у вигляді жовткових включень, рівномірно розподілених у клітині. Ззовні яйцеклітина оточена оболонками: жовтковою, прозорою і зовнішньою, що складається з фолікулярних клітин і виконує функції живлення та захисту.
Мал.2. Будова яйцеклітини
Складний процес розмноження значною мірою залежить від розвитку статевих клітин, їхньої функціональної (репродуктивної) здатності.
Органи чоловічої статевої системи.
Підрозділяється на внутрішні та зовнішні статеві органи. До внутрішніх чоловічих статевих органів відносяться: статеві залози — яєчка, або сім'яники (testis), які покриті щільною сполучнотканинною оболонкою і містять канальці загальною довжиною до 300-400 м., в яких протягом усього життя утворюються сперма тозоїди; придатки яєчка, де нагромаджуються зрілі сперматозоїди; сім’яні пухирці; передміхурова (prostata) і куперова (glandula bulbourethralis) залози, що утворюють секрети, створюючи певне хімічне середовище для сперматозоїдів. Сперматозоїди разом із секретом додаткових залоз являють собою сперму. Зовнішні статеві органи включають мошонку (scrotum), в якій знаходяться яєчка та їх придатки, і статевий член, або пеніс (penis), що служить для введення сперми в статеві шляхи жінки. Мошонка є випинанням стінки тіла, куди спускаються яєчка напередодні або незабаром після народження.
Р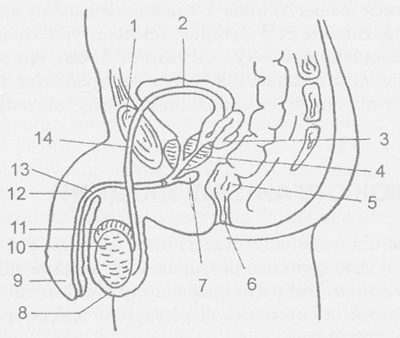 ис.
102. Чоловіча статева система: 1— лобкова
кістка; 2— сечовий міхур; З — сім'яний
пухирець; 4 — передміхурова залоза; 5 —
пряма кишка; 6 — анальний отвір; 7 —
куперова залоза; 8 — мошонка; 9 — голівка
пеніса; 10 — сім'яник; 11 — придаток
сім'яника; 12 — пеніс; 13 — сечовипускальний
канал; 14 — сім'явиносна протока.
ис.
102. Чоловіча статева система: 1— лобкова
кістка; 2— сечовий міхур; З — сім'яний
пухирець; 4 — передміхурова залоза; 5 —
пряма кишка; 6 — анальний отвір; 7 —
куперова залоза; 8 — мошонка; 9 — голівка
пеніса; 10 — сім'яник; 11 — придаток
сім'яника; 12 — пеніс; 13 — сечовипускальний
канал; 14 — сім'явиносна протока.
Внутрішні статеві органи виконують ендокринні функції. В канальцях сім'яників крім сперматогенного епітелію знаходяться так звані підтримуючі й інтерстиціальні клітини, одна з функцій яких — утворення чоловічого статевого гормону — тестостерону. Передміхурова залоза секретує гормони, регулюючі обмін речовин у клітинах — простагландини.
Органи жіночої статевої системи.
Ж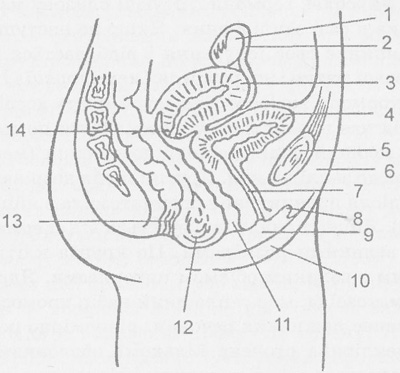 іноча
статева система представлена внутрішніми
та зовнішніми статевими органами.
Внутрішні статеві органи — це
яєчники,яйцепроводи, матка і піхва, які
розташовані в малому тазу. Зовнішні
статеві органи — великі, малі статеві
губи і клітор.
іноча
статева система представлена внутрішніми
та зовнішніми статевими органами.
Внутрішні статеві органи — це
яєчники,яйцепроводи, матка і піхва, які
розташовані в малому тазу. Зовнішні
статеві органи — великі, малі статеві
губи і клітор.
Рис. 103. Жіноча статева система: 1 — яєчник; 2 — яйцепровід; 3 — матка; 4 — сечовий міхур; 5 — лобкова кістка; 6 — лобок; 7 — сечовипускальний канал; 8 — клітор; 9 — мала статева губа; 10 — велика статева губа; 11 — піхва; 12 — м'язи промежини; 13— пряма кишка; 14 — шийка матки.
Яєчники (статеві залози) (ovarium) — це невеликі залози завдовжки 3-4 см., масою 6-7 гр. Вони містять овоцити першого порядку, котрі оточені шаром фолікулярних клітин. Така структура називається фолікулом. З овоцитів першого порядку утворюються жіночі статеві клітини — яйцеклітини. У міру дозрівання яйцеклітини, фолікул наближається до поверхні яєчника, у порожнині його нагромаджується рідина, внаслідок чого фолікул розривається і яйцеклітина потрапляє в яйцепровід (oviductus) і потім у матку (uterus). Матка є порожнистим гладком'язевим органом, що складається зі дна, тіла і шийки. Тіло складає велику частину матки і, звужуючись униз, переходить у шийку. Шийка матки вдається у верхню частину піхви. При вагітності матка збільшується і змінює свою форму. Піхва (vagina) — м'язова трубка, що своїм верхнім кінцем охоплює шийку матки, а нижнім відкривається в статеву щілину.
З періоду статевого дозрівання яєчники починають виробляти й виділяти жіночі статеві гормони, тобто яєчники починають функціонувати і як залози внутрішньої секреції. Розвиток яйцеклітин починається з періоду статевого дозрівання (12-13 років) і відбувається періодично один раз на місяць до 45-50 років, коли ендокринна функція яєчників згасає. На місці фолікула, що лопнув, розвивається тимчасова залоза внутрішньої секреції — жовте тіло, яка виробляє гормони, готуючи слизову матки для прийняття зародка в разі запліднення. Якщо не наступає вагітності, ця залоза припиняє своє існування і відбувається відторгнення епітелію слизової матки, що розрісся (менструація). При настанні вагітності гормони жовтого тіла затримують дозрівання інших фолікулів. Під час проходження яйцеклітини по матковій трубі закінчується перший поділ періоду дозрівання (мейоз-1). Яйцеклітина готова до запліднення. Другий поділ дозрівання (мейоз-2) починається після проникнення сперматозоїда в яйцеклітину (активація яйця). Яйцеклітина відрізняється від сперматозоїда будовою і більш великими розмірами. Це кругла клітина розміром близько 0,1 мм. з великим об'ємом цитоплазми. Ядро яйцеклітини, як і сперматозоїда, має гаплоїдний набір хромосом. Цитоплазма містить запас поживних речовин, рівномірно розподілених у клітині. Яйцеклітина оточена кількома оболонками. Зовнішня оболонка складається з фолікулярних клітин, виконуючих поживну та захисну функції.
Будова зрілих статевих клітин.
На відміну від соматичних, статеві клітини (гамети – від слова “молодята”) спеціалізовані на відтворенні поколінь організмів і мають половинний (гаплоїдний) генетичний набір (lcln, або 23 хромосоми – у людини).
Розрізняють чоловічі і жіночі статеві клітини, які несуть генетичну інформацію по батьківській і материнській лініях. У статевих клітинах у людини присутні 22 аутосоми і 1 статева хромосома, яка позначається як X або Y – у чоловіків і X – у жінок. При злитті чоловічої та жіночої статевих клітин в процесі запліднення утворюється диплоидная клітина – зигота, яка дає початок усім клітинам нового організму. В ембріогенезі людини статеві клітини вперше виявляються в желточной ентодермі, тобто внегонаднимі, а потім мігрують у закладку статевих залоз.
При першому знайомстві з будовою зрілих чоловічих і жіночих статевих клітин, звертає на себе увагу їх висока функціональна доцільність. Сперматозоїди, які повинні бути високомобільні в жіночому статевому тракті, являють собою маленькі клітини, практично позбавлені цитоплазми і складаються з ядерні частини, або головки, що несе генетичний матеріал, і органу пересування – хвоста, або джгутика. Ніяких інших субклітинних елементів, крім мітохондрій, що дають енергію для пересування, акросомальної вакуолі з протеолітичними ферментами для розчинення оболонок яйцеклітини, і проксимальної центріолі, сперматозоїди не мають. Загальна довжина спермія становить близько 60 мкм, з яких на частку хвоста припадає 55 мкм.
На ультрамікроскопічні рівні в головці сперматозоїда помітні акросомальная і постакросомальная зони, в хвості – проміжний, головний і кінцевий відділи. Велику частину головки займає ядро, заповнене конденсованим хроматином. Простір між ядром і передньою ділянкою плазмолемми спермія зайнято акросомальної вакуолью – спеціалізованої лізосомах, що містить групу ферментів-лізину оболонок яйцеклітини: акрозін (руйнує прозору зону овоцитів), пенетраза (диссоциирует клітини променистого вінця), гіалуронідаза (розщеплює гіалуронову кислоту), кисла фосфатаза (руйнує форсхолін при проходженні сперміїв через плазмолему овоцитів). Шийка спермія являє собою короткий відділ, в якому знаходяться проксимальна центриоль і 9 сегментованих колон.
Проміжний відділ містить аксонів і 9 поздовжньо орієнтованих елементів цитоскелета, що складаються з кератіноподобним білків і є продовженням сегментованих колон. Назовні від цих волокон розташовуються мітохондрії. Аксонема побудована за принципом вії, биття якої ініціюються катіонами кальцію, що знаходяться в навколишньому середовищі, і мітохондріальної АТФ спермія. В кінцевому відділі хвоста елементи цитоскелета зникають, і аксонема прикрита лише плазмолеммой. У людини дуплети аксонема в кінцевому відділі розпадаються на 18 одиночних мікротрубочок.
Після виходу з яєчка спермін морфологічно сформовані, але функціонально незрілі – вони нерухомі, не здатні до запліднення і, крім цього, є носіями низки специфічних антигенів, інактивація яких (як і придбання рухливості і запліднюючої здібності – капацитации) відбувається в системі сім’явиносних шляхів чоловічого статевого тракту.
Яйцеклітина на відміну від сперматозоїдів велика й нерухома. У більшості ссавців і людини її розміри сягають 100-200 мкм. Жіноча статева клітина є берегинею запасу трофічних сполук, необхідних для раннього розвитку зародка, і “запасу” цитоплазматичних структур для формування перших генерацій зародкових клітин – бластомерів. У людини великі розміри яйцеклітини не пов’язані з великими запасами жовтка – яйцеклітина людини бідна жовтком (Олиголецитальная).
На ультрамікроскопічні рівні яйцеклітина людини має велике округле ядро, в якому переважає еухроматин, і цитоплазму з помірною кількістю мітохондрій з нечисленними ламеллярние кристами. Добре розвинена система білкового синтезу через високий вміст рибо-і полісом, коротких канальців гранулярної ендоплазматичної мережі. В цитоплазмі рідко і дифузно розташовані нечисленні скупчення вітеллінових гранул – колекторів трофічних включень.
Слід особливо підкреслити, що у вищих плацентарних ссавців і людини жіночі статеві клітини не існують ізольовано. За винятком самих ранніх стадій розвитку (первинних статевих клітин, овогоній) вони знаходяться в тісному контакті з соматичними клітинами яєчника (фолікулярними епітеліоцитами і сполучнотканинними клітинами), які формують навколо кожної статевої клітини епітеліальну ісполучнотканинних оболонки. Комплекс “статева клітина – соматичні оболонки” іменується оваріальних фолікулом, або ово-соматичним (фолікулярним) гістіоні. Будова оваріального фолікула ускладнюється в процесі овогенезу.
Гаметогенез, його періоди.
Гаметогенез — розвиток статевих клітин — гамет— чоловічих (сперматозоїдів) і жіночих (яйцеклітин) відбувається в статевих залозах: сперматогенез — у чоловічих статевих залозах (сім’яниках, яєчках), овогенез — у жіночих статевих залозах (яєчниках).
Гаметогенез проходить такі стадії: розмноження, росту, дозрівання статевих клітин, а сперматогенез проходить ще окрему четверту стадію — формування. Мейоз відбувається в період дозрівання статевих клітин. Вступає в перший поділ мейозу гаметоцит (спермато- чи овоцит) першого порядку, клітина, яка має диплоїдний набір (у людини 46) (d-хромосом. У результаті двох поділів дозрівання (мейозу) утворюються клітини з гаплоїдним набором хромосом (у людини 23 s-хромосоми). У чоловіків на стадії формування зріла статева клітина набуває остаточної форми — сперматозоїда з головкою і хвостом. Формування жіночої статевої клітини (оотиди) відбувається одночасно з ростом і дозріванням і полягає в накопиченні жовтка і утворенні оболонки яйцеклітини.
Сперматогенез та овогенез.
Оогенез — процес розвитку жіночих статевих клітин і оогенної тканини. Оогенез проходить у яєчниках у три періоди: 1) розмноження (клітини диплоїдної оогенної тканини діляться шляхом мітозу, утворюючи диплоїдні ооцити); 2) ріст ооцитів, проходження ними клітинного циклу, під час якого відбувається синтез ДНК, подвоєння і побудова другої хроматиди хромосом; 3) дозрівання ооцитів (їх поділ шляхом мейозу). У результаті оогнезу з ооцита утворюється одна гаплоїдна яйцеклітина з однохроматидними хромосомами і три редукційних (чи полярних) тільця. Далі яйцеклітина бере участь у статевому процесі, а редукційні тільця відмирають.
Оогенез
С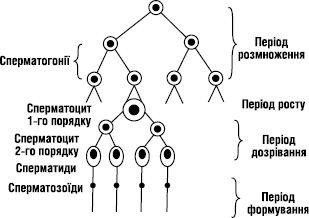
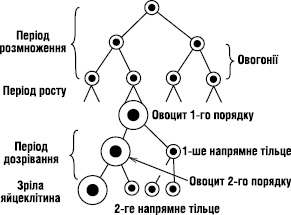 перматогенез
(лат. spermato — сім’я + genesis — походження)
— процес розвитку статевих клітин —
сперматозоїдів — зі сперматогенної
тканини. Сперматогенез проходить у три
стадії: 1) розмноження в сім’яниках
клітин диплоїдної сперматогенної
тканини, що поділяються шляхом мітозу
та утворюють диплоїдні сперматоцити;
2) ріст сперматоцитів, проходження ними
клітинного циклу, у ході якого здійснюються
синтез ДНК і добудування другої хроматиди;
3) дозрівання сперматоцитів, що поділяються
шляхом мейозу, в результаті чого
утворюються гаплоїдні сперматозоїди.
З кожного сперматоцита виникає по чотири
гаплоїдні однохроматидні сперматозоїди,
готові до запліднення. При цьому в
кожному сперматозоїді (людини та інших
ссавців) хромосомні набори розрізняються
за статевими хромосомами: одні несуть
Х-хромосому, інші — Y-хромосому, а всі
хромосоми несуть різні комбінації
генів, набуті в результаті обміну
ділянками у I профазі мейозу.
перматогенез
(лат. spermato — сім’я + genesis — походження)
— процес розвитку статевих клітин —
сперматозоїдів — зі сперматогенної
тканини. Сперматогенез проходить у три
стадії: 1) розмноження в сім’яниках
клітин диплоїдної сперматогенної
тканини, що поділяються шляхом мітозу
та утворюють диплоїдні сперматоцити;
2) ріст сперматоцитів, проходження ними
клітинного циклу, у ході якого здійснюються
синтез ДНК і добудування другої хроматиди;
3) дозрівання сперматоцитів, що поділяються
шляхом мейозу, в результаті чого
утворюються гаплоїдні сперматозоїди.
З кожного сперматоцита виникає по чотири
гаплоїдні однохроматидні сперматозоїди,
готові до запліднення. При цьому в
кожному сперматозоїді (людини та інших
ссавців) хромосомні набори розрізняються
за статевими хромосомами: одні несуть
Х-хромосому, інші — Y-хромосому, а всі
хромосоми несуть різні комбінації
генів, набуті в результаті обміну
ділянками у I профазі мейозу.
Сперматогенез
Мейоз.
Мейоз (або редукційний поділ) — особливий вид поділу еукаріотичнихклітин, характерний тільки статевимклітинам (не соматичним), унаслідок якого хромосомний набір зменшується вдвічі, клітини переходять з диплоїдногостану в гаплоїдний.
Мейоз складається з двох послідовних поділів, аналогічних мітотичним (з деякими відмінностями), інтерфаза між якими вкорочена, а у рослинних клітинах може бути взагалі відсутня
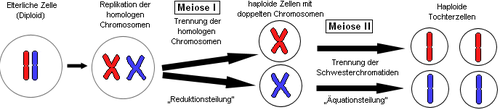
Стадії мейозу.
Перший поділ мейозу. У профазі-І виділяють таю стадії: лептотенна, зиготенна, пахітенна, диплотенна і діакінез.
1) Лептотенна стадія, або лептотена (від грецьк. leptos — тонкий, дрібний і teino — розтягаю), або лептонема (від грецьк. nema — нитка), характерна тим, що в ній хромосоми починають конденсуватися, але мають вигляд тонких ниток, тому на них може синтезуватися певна кількість РНК. Ядерце і ядерна оболонка (каріолема) зберігаються. Цей період деякі автори називають пролептонемою (від грецьк. рго — період). Відтак спіралізація хромосом триває. Кожна з 46 (d-хромосом за допомогою прикріплювальних дисків приєднується обома кінцями до внутрішньої мембрани каріолеми.
Зиготенна стадія, або зиготена (від грецьк. zygon — пара), названа так тому, що під час її перебігу гомологічні хромосоми наближаються одна до одної, розташовуються парами, вкорочуються, обвиваються, з’єднуються між собою. У місцях контактів.гомологічних хромосом утворюються своєрідні білкові структури — синаптонемні комплекси (від грецьк. synapsis — зв’язок, з’єднання), завдяки яким названі хромосоми кон’югують (від латин, conjungo — з’єднувати), формуючи біваленти (від латин. bi — подвійний і valens — сильний). Нагадаємо, що кожна диплоїдна хромосома з одного бівалента походить або від батька, або від матері (46 (d-хромосом утворюють 23 біваленти, або тетради хроматид). Статеві хромосоми розташовуються біля внутрішньої ядерної мембрани в ділянці, названій статевим міхурцем. Гетерохромосоми X і Y кон’югують неповністю, оскільки вони не зовсім гомологічні.
1) Пахїтенна стадія, або пахітена (від грецьк. pachys — товстий, повний) при гаметогенезі триває щонайменше кілька діб. Хромосоми поступово вкорочуються і потовщуються. Між хроматидами материнського і батьківського походження у декількох місцях виникають хіазми (від грецьк. chiasma — перехрест), або рекомбінантні вузлики, у вигляді білкових комплексів розміром приблизно 90 нм. У ділянках кожної хіазми відбувається обмін відповідними ділянками гомологічних хромосом — кросинговер (від англ. crossing і over — перехрест). Цей процес забезпечує численні генетичні рекомбінації. У кожному бівалєнті людини кросинговер відбувається у двох-трьох ділянках, що відповідає кількості рекомбінантних вузликів.
2) Диплотенна стадія, або диплотена (від грецьк. diploos — подвійний), характерна тим, що синаптонемні комплекси розпадаються, кон’югованіхромосоми розсуваються і гомологічні хромосоми відходять із бівалентів, зберігаючи зв’язок лише у місцях хіазм. Ця стадія при гаметогенезі досить тривала і поступово переходить у наступну.
3) У діакінезі (від грець, (dia — через і kinesis — рух) хромосомипродовжують потовщуватися, відділяються від нуклеолеми. При цьому гомологічні (d-хромосоми продовжують залишатися зв’язаними між собою хіазмами, а сестринські хроматиди кожної d-хромосоми — центромерами. Завдяки наявності кількох хіазм біваленти утворюють петлі. У цей час руйнується ядерна оболонка і ядерця. Репліковані центріолі спрямовуються до полюсів.
Запліднення та його фази.
ЗАПЛІДНЕННЯ (лат. fertilisatio) — злиття чоловічої та жіночої статевих клітин, унаслідок чого відновлюється диплоїдний набір хромосом, характерний для даного виду, та утворюється якісно нова клітина — зигота (запліднена яйцеклітина, або одноклітинний зародок). У людини об’єм еякуляту — викинутої сперми — у нормі становить близько 3 мл. Для забезпечення З. загальна кількість сперматозоїдів у спермі повинна становити не менше 150 млн, а їх концентрація в 1 мл — 20–200 млн, хоча в яйцеклітину проникає тільки один з них, а інші готують умови для З. У статевих шляхах жінки після копуляції їх кількість зменшується в напрямку від піхви до дистального кінця маткової труби, куди яйцеклітина надходить з черевної порожнини, зберігаючи здатність до З. протягом 2–3 діб. У процесі З. розрізняють 3 фази:
1) дистантна взаємодія та зближення гамет;
2) контактна взаємодія й активізація яйцеклітини;
3) входження сперматозоїда в яйцеклітину та наступне злиття — сингамія.
Перша фаза — дистантна взаємодія — забезпечується хемотаксисом — сукупністю специфічних факторів, які підвищують імовірність зіткнення статевих клітин. Важливу роль у цьому відіграють гормони — хімічні речовини, які виробляються статевими клітинами. Встановлено, що яйцеклітини виділяють пептиди, які сприяють залученню сперматозоїдів. Відразу після еякуляції сперматозоїди не здатні проникнути в яйцеклітину доти, поки не відбудеться капацитація — набуття сперматозоїдами запліднювальної здатності під дією секрету жіночих статевих шляхів, що триває 7 год. У процесі капацитації з плазмолеми сперматозоїда в ділянці акросоми віддаляються глікопротеїни, які сприяють акросомальній реакції. У механізмі капацитації велике значення належить гормональним факторам, насамперед прогестерону (гормон жовтого тіла), що активізує секрецію залозистих клітин яйцепроводів. Під час капацитації відбувається зв’язування холестерину цитолеми спермія альбумінами жіночих статевих шляхів та оголення рецепторів статевих клітин. З. відбувається в ампулярній частині маткової труби.
Перед З. відбувається взаємодія та зближення гамет (дистантна взаємодія), зумовлене хемотаксисом. Друга фаза З. — контактна взаємодія, під час якої сперматозоїди обертають яйцеклітину. Численні сперматозоїди наближаються до яйцеклітини та вступають у контакт із її оболонкою. Яйцеклітина починає робити обертальні рухи навколо своєї осі зі швидкістю 4 оберти за хвилину. Ці рухи зумовлені биттям джгутиків сперматозоїдів та продовжуються близько 12 год. У процесі взаємодії чоловічої та жіночої статевих клітин у сперматозоїдах відбувається акросомальна реакція. Вона полягає в злитті зовнішньої мембрани акросоми з передніми 2/3 плазмолеми сперматозоїда. Потім у ділянці злиття мембрани розриваються, і ферменти акросоми виходять у навколишнє середовище. Ініціація другої фази З. відбувається під впливом сульфатованих полісахаридів блискучої зони, що викликають надходження кальцію і натрію в голівку чоловічої гамети, заміщення ними іонів калію і водню та розриву мембрани акросоми. Прикріплення сперматозоїда до яйцеклітини відбувається під впливом вуглеводної групи фракції глікопротеїнів прозорої зони яйцеклітини. Рецептори сперматозоїда для прозорої зони — фермент глікозилтрансфераза, який знаходиться на поверхні акросоми голівки, що «розпізнає» цукор N-ацетилглюкозамін — рецептор жіночої статевої клітини. Плазматичні мембрани в місці контакту статевих клітин зливаються й відбувається плазмогамія — об’єднання цитоплазм обох гамет. Сперматозоїди при контакті з яйцеклітиною можуть зв’язувати десятки тисяч молекул глікопротеїду Zp3. При цьому запускається акросомальна реакція. Ця реакція характеризується підвищенням проникності плазмолеми спермія до іонів Са2+, її деполяризацією, що сприяє злиттю плазмолеми з передньою мембраною акросоми. Блискуча зона виявляється в безпосередньому контакті з акросомальними ферментами. Ферменти руйнують блискучу зону, спермій проходить крізь неї та входить у перивітеліновий простір, розташований між блискучою зоною та плазмолемою яйцеклітини. Через кілька секунд змінюються властивості плазмолеми яйцеклітини, починається кортикальна реакція, а через кілька хвилин змінюються властивості блискучої зони — Zp (зонна реакція) (рисунок).
Р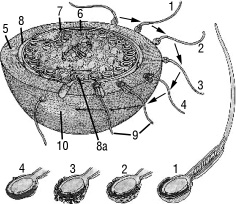 исунок.
З.: 1, 2, 3, 4 — стадії акросомальної реакції;
5 — блискуча зона (zona pellucida); 6 —
перивітеленовий простір; 7 — плазматична
мембрана; 8 — кортикальна гранула; 8а —
кортикальна реакція; 9 — входження
спермія в яйцеклітину; 10 — зонна реакція
исунок.
З.: 1, 2, 3, 4 — стадії акросомальної реакції;
5 — блискуча зона (zona pellucida); 6 —
перивітеленовий простір; 7 — плазматична
мембрана; 8 — кортикальна гранула; 8а —
кортикальна реакція; 9 — входження
спермія в яйцеклітину; 10 — зонна реакція
У ссавців при З. в яйцеклітину проникає лише один сперматозоїд. Таке явище називається моноспермією. З. сприяють сотні інших сперматозоїдів, які беруть участь у З. Ферменти, що виділяються з акросом, — спермолізини (трипсин, гіалуронідаза) — руйнують променистий вінець, розщеплюють глікозаміноглікани прозорої зони яйцеклітини. Фолікулярні клітини, які відокремлюються, склеюються в конгломерат, який слідом за яйцеклітиною переміщається по трубі завдяки мерехтінню війок епітеліальних клітин слизової оболонки. У третій фазі в ооплазму проникають голівка і проміжна частина хвостового відділу. Після входження сперматозоїда в яйцеклітину на периферії ооплазми відбувається її ущільнення (зонна реакція) та утворюється оболонка З. Кортикальна реакція — злиття плазмолеми яйцеклітини з мембранами кортикальних гранул, унаслідок чого вміст із гранул виходить у перивітеліновий простір і впливає на молекули глікопротеїдів блискучої зони. Внаслідок цієї зонної реакції молекули Zp3 модифікуються та втрачають здатність бути рецепторами сперматозоїдів. Утворюється оболонка З. товщиною 50 нм, яка перешкоджає поліспермії — проникненню інших сперміїв. Механізм кортикальної реакції включає надходження іонів натрію крізь ділянку мембрани сперматозоїда. Внаслідок цього негативний мембранний потенціал клітини стає слабкопозитивним. Надходження іонів натрію зумовлює вивільнення іонів кальцію із внутрішньоклітинних депо та збільшення його вмісту в гіалоплазмі яйцеклітини. Після цього починається екзоцитоз кортикальних гранул. З них вивільняються протеолітичні ферменти, які розривають зв’язки між блискучою зоною та плазмолемою яйцеклітини, а також між сперматозоїдами та прозорою зоною. Крім того, виділяється глікопротеїд, що зв’язує воду та залучає її в простір між плазмолемою і блискучою зоною. Унаслідок цього формується перивітеліновий простір. Нарешті, виділяється фактор, що сприяє затвердінню прозорої зони й утворенню з неї оболонки З. Завдяки механізмам запобігання поліспермії тільки одне гаплоїдне ядро сперматозоїда одержує можливість злитися з одним гаплоїдним ядром яйцеклітини, що приводить до відновлення характерного для всіх клітин диплоїдного набору. Проникнення сперматозоїда в яйцеклітину через кілька хвилин значно підсилює процеси внутрішньоклітинного обміну, що пов’язано з активізацією її ферментативних систем. Взаємодія сперматозоїдів з яйцеклітиною може бути заблокована за допомогою антитіл до речовин, що входять у прозору зону. На цій основі розробляють засоби імунологічної контрацепції. Після зближення жіночого та чоловічого пронуклеусів, що продовжується у ссавців приблизно 12 год, утворюється зигота — одноклітинний зародок. Уже на стадії зиготи виявляються презумптивні зони як джерела розвитку відповідних ділянок бластули, з котрих надалі формуються зародкові листки. У людини в процесі овуляції з яєчника звичайно виділяється одна яйцеклітина. Тому після З. утворюється одна зигота, з якої розвивається, як правило, один плід. Рідко у процесі дроблення зиготи на ранніх стадіях відбувається відокремлення бластомерів. У цих випадках кожний з них дає початок окремому організму — народжуються монозиготні (однояйцеві) близнюки, генетично цілком ідентичні. Іноді при овуляції з яєчника виділяються дві і більше яйцеклітини, кожна з яких може бути запліднена. У таких випадках з кожної зиготи формується плід, що має генетичні розбіжності з іншими плодами. Це дизиготні (різнояйцеві) близнюки, що нерідко є різностатевими. Використання контрацептивних препаратів запобігає З., зумовлюючи ановуляторний менструально-оваріальний цикл (естроген-гестогенні препарати для систематичного застосування) чи пригнічення життєдіяльності сперматозоїдів (посткоїтальні препарати). Лікування безпліддя, навпаки, спрямоване на стимуляцію гаметогенезу, овуляції, З., а також подальше виношування вагітності.
Дроблення, його типи, фактор. що зумовлюють тип дроблення.
Після запліднення яйцеклітина починає ділитися. Процес ділення яйцеклітини, що приводить до збільшення числа клітин без їхнього росту, називають дробленням. Характер дроблення визначається кількістю жовтка в яйцеклітині і його розподілом. В залежності від цього дроблення може бути повним і частковим .. Дроблення, при якому яйцеклітина ділиться повністю, називають голобластіческімі, а неповністю, частково - меробластіческімі.
Повний дроблення яйцеклітини може бути рівномірним і нерівномірним.
Повне рівномірне дроблення властиво яйцеклітинам ланцетника, яйця якого мають мало жовтка. При цьому виді дроблення клітини, які утворюються, або бласто заходи, мають приблизно однакові розміри.
Повний нерівно мірне дроблення властиво яйцеклітинам хрящових риб, земноводних, яйця яких мають більше жовтка. При цьому виді дроблення утворюються неоднакові за величиною бластомери, причому перші два дроблення дають рівні бластомери, а потім бластомери, розташовані на анімальному полюсі, діляться швидше, стають дрібнішими, ніж на вегетативному.
Неповне дроблення яйцеклітин може бути поверхневим і дискоидальная.
При неповному поверхневому дробленні, яке притаманне багатьом членистоногим, дробиться лише поверхнева частина яйцеклітини, а центральна її частина, багата жовтком, не ділиться.
При неповному дискоидальная дроблю нии, яке властиво, наприклад, гадів, птахам, дробиться лише невелику ділянку поверхні яйцеклітини, де мало жовтка, а інша її частина, багата жовтком, не ділиться. У зв'язку з тим що при цьому виді дроблення поверхневий ділянку роздрібнюється яйця має форму диска, то і дроблення отримало найменування дискоидальная (рис. 1).
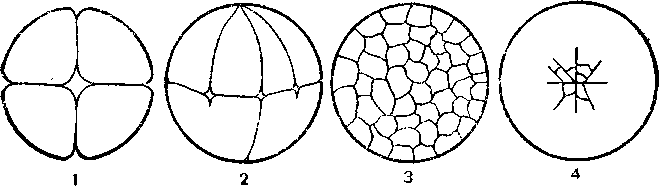 Рис.
1. Типи дроблення яйцеклітин (оріг.):
Рис.
1. Типи дроблення яйцеклітин (оріг.):
1 - повне рівномірне; 2 - повне нерівномірне; 3 - неповне поверхневе; 4 - неповне діскондальное.
Характер дроблення визначається не тільки кількістю жовтка і розподілом його в яйцеклітині, але і взаємним розташуванням клітин, які утворюються в результаті дроблення.
По напрямку дроблення розрізняють радіальне, спіральне, двусімметрічное, або білатерально, дроблення.
При радіальному дробленні, яке властиво кишечнополостним, голкошкірі і багатьом нижчим хордових, верхній ряд бластомерів розташовується точно над нижнім.
При спіральному дробленні, яке спостерігається у більшості черв'яків і молюсків, верхній ряд бластомерів розташовується між бластомерами нижнього ряду.
При білатеральному дробленні, яке властиво круглим черв'яки, асцидії, бластомери розташовуються симетрично з боків вихідного бластомера. Слід зауважити, що в процесі дроблення яйцеклітини часто має місце поєднання різних типів дроблення.
Бластула, морфологічні особливості різних видів бластул.
В процесі дроблення виникає багатоклітинний одношаровий зародок, який зовні нагадує кулю, у багатьох випадках з порожниною в середині. Такий зародок називають бластулою. Шар клітин, який утворює стінку бластули, називають бластодермой (Derma - шкіра), а порожнина бластули - бластоцелем (Koilon - порожнина) або первинної порожниною тіла. Коли ж при дробленні виникає кулястий зародок без порожнини всередині, схожий на ягоду шовковиці, то його називають морулой (Morula ягода шовковиці). Така бластула властива, наприклад, плацентарних ссавців.
В залежності від типу дроблення яйцеклітини розрізняють, крім морули, ще 5 типів бластул: целобластула, амфібластула, стерробластулу, перібластулу і діскобластулу.
Бластули, бластодерма якої утворена одним шаром клітин, а бластоцель займає центральне положення, називають типовою бластулою або целобластула. Така бластула властива, наприклад, ланцетнику.
Амфібластула відрізняється від типової бластули тим, що її бластоцель зміщений до анімальному полюсі - зоні дрібних бластомерів, а бластодерма представлена кількома рядами клітин. Така бластула властива земноводним, від яких і отримала своє найменування.
Стерробластула (Sterros - щільний) складається з великих клітин бластодерми, які настільки глибоко заходять в порожнину бластоцелем, що від останнього нічого не залишається. Таку бластули мають деякі членистоногі.
У перібластуле, що виникає при поверхневому дробленні яйця, клітини бластодерми розташовуються по периферії недробящегося жовтка і бластоцелем немає. Такий тип бластули властивий деяким комахам.
Дискоїдальне дроблення веде до формування діскобластули, бластоцель якої має вигляд вузької щілини, розташованої під клітками бластодерми, іменованої в даному випадку зародковим диском. Такий тип бластули спостерігається у костистих риб, плазунів, птахів.
Р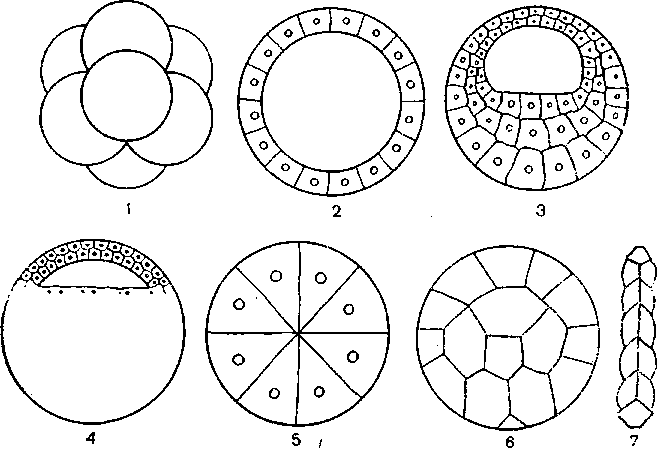 ис.
2. Типи бластул (оріг.):
ис.
2. Типи бластул (оріг.):
/ - Морула; 2 - цслобластула; 3 - амфнбластула; 4 - діскобластула; 5 - стерробластула; 6 - перібластула; 7 - плакула.
Розглянуте різноманіття бластул показує, що на цій стадії розвитку організмів має місце диференціювання клітинного матеріалу зародка, пов'язана з кількістю і розташуванням жовтка, з вмістом і синтезом ДНК, з поділом речовин у цитоплазмі клітин.
На процеси дроблення і ранні стадії розвитку організмів впливають умови зовнішнього середовища,оскільки розвиток організму відбувається в середовищі, в якому проходило його формування в історичному розвитку. Незначні відхилення умов середовища від типових позначаються в основному лише на темпах дроблення.
До факторів зовнішнього середовища, які впливають на дроблення яйцеклітини і ранні стадії розвитку організму, слід в першу чергу віднести наявність вологи, хімічний склад і реакцію середовища (рН), світловий потік, температуру, наявність поживних речовин, кисню та ін
Гаструляція, способи гаструляції.
Гаструляція - це процес утворення двуслойного зародка.
У більш широкому поданні г аструляція - складний процес хімічних і морфологічних змін, що супроводжується розмноженням, ростом, спрямованим переміщенням і диференціюванням клітин, в результаті чого утворюються зародкові листки, джерела зачатків тканин і органів, комплекси осьових органів.
Гаструляція у більшості тварин відбувається в два етапи. На першому етапі утворюються 2 зародкових листка (ектодерма і ентодерми), а на другому формується третій 3 зародковий листок (мезодерма) і відбуваються подальше формування ембріона.
Розрізняють чотири типи, або способу, утворення двуслойного зародка: вселення, чи імміграція; впячивание, або інвагінація; обростання, або епіболія; розшарування, або деляминации.
Вихідною в процесі філогенетичного розвитку і найбільш примітивною формою гаструляціі є вселення, чиімміграція. Такий спосіб утворення двуслойного зародка полягає в тому, що окремі клітини або групи клітин активно переміщаються з бластодерми в бластоцель. Ці клітини, що виникають внаслідок поділу клітин бластодерми, осідають на її внутрішній поверхні, утворюючи внутрішній шар клітин, або ентодерми, а клітини бластодерми, розташовані зовні, іменуються вже ектодерми. Якщо вселення клітин в бластоцель має місце лише з боку одного вегетативного полюса бластули, то така імміграція носить назву уніполярної або однополюсної, а коли з різних ділянок бластодерми, - мультиполярної або багатополюсної. Уніполярна імміграція властива деяким гідроїдних поліпів, медуз і майже всім гідромедуз. Мультиполярні імміграція є рідкісним явищем і спостерігається у деяких медуз. При імміграції гастраль ная порожнину, або гастроцель(Порожнина первинної кишки), утворюється або одночасно з виникненням двуслойного зародка, як це має місце, наприклад, у гідромедуз, або значно пізніше, як у багатьох видів кишковопорожнинних. У останніх спостерігається настільки масове вселення клітин, що бластоцель ними повністю заповнюється, порожнину зникає і лише згодом, з відокремленням ентодерми, виникає гастроцель, природно, що не має зв'язку із зовнішнім середовищем. Цей спосіб гаструляції особливо поширений серед двухшарових тварин - кишечнополостних. Всі інші способи гаструляції є похідними імміграції.
Якщо гаструляция здійснюється шляхом впячивания, або інвагінації (In-проникнення, vagina - оболонка, піхву), то одношарова стінка бластули - бластодерма - прогинається всередину бластоцелем і досягає протилежної стінки. Впячивание починається з вегетативного полюса, який розпізнається за більш великим клітинам, і йде в напрямку анімального полюса. По закінченні інвагінації зародок стає двошаровий, оскільки бластодерма утворює два шари клітин: зовнішній - ектодерму і внутрішній - ентодерми. Після впячиванія формується порожнина, яка оточена клітинами ентодерми. Цю порожнину називають гастрального порожниною. Гастроцель повідомляється із зовнішнім середовищем отвором, який називають первинним ротом або бластопором). Краю цього отвору іменуються губами.
У процесі еволюційного розвитку представників тваринного царства формування бластопор при гаструляції послужило підставою для поділу тварин на дві великі групи за такою ознакою, як освіта первинного рота.У хробаків, молюсків, членистоногих первинний рот, розвиваючись і диференціюючись, перетворюється в постійний, або дефінітивний, рот дорослої особини. Таких тварин називають первичноротих. У голкошкірих, кишководишних, хордових бластопор перетворюється в заднепроходное, або анальний отвір, або в нервово-кишковий канал, який знаходиться на задньому кінці ембріона, а ротовий отвір виникає заново на передньому кінці черевної поверхні зародка. Таких тварин іменують Вториннороті. Гаструляция шляхом впячиванія дуже поширена у нижчих Вториннороті і хордових.
Доля бластоцелем при інвагінації може бути двоякою: він може зберегтися в тому випадку, якщо клітинні шари ектодерми і ентодерми не стикаються між собою і тоді у зародка спостерігаються одночасно дві порожнини - бластоцель і гастроцель. Якщо ж клітинні шари ектодерми і ентодерми при впячивание стикаються на всьому протязі, то бластоцель зникає і у зародка спостерігається лише одна порожнина - порожнина первинної кишки, або гастроцель.
У деяких хребетних тварин у зв'язку зі збільшенням кількості жовтка в яйцеклітині і незначними розмірами бластоцелем, відтісняються до анимального полюси, гаструляция лише за типом інвагінації стає неможливою. Тоді у деяких круглоротих і земноводних до впячивание приєднується обростання, абоепіболія. Гаструляція шляхом епіболіі полягає в тому, що клітини бластодерми анімального полюса бластули діляться значно швидше, ніж клітини вегетативного полюса. У зв'язку з цим дрібні клітини анімального полюса поступово зовні наповзають на великі клітини вегетативного полюса, обростаючи їх і утворюючи ектодерму, а клітини вегетативного полюса формують ентодерми. При гаструляціі шляхом епіболіі в чистому вигляді, як це має місце у деяких безхребетних, спочатку немає ні бластопор, ні гастроцель, які виникають значно пізніше. У тих же випадках, коли епіболія супроводжує інвагінації, як це спостерігається у деяких костистих риб, земноводних, то виникає бластопор, а також порожнину первинної кишки, які існують поряд з бластоцелем.
У костистих риб, плазунів, птахів і яйцекладущих ссавців, яйцеклітини яких мають дуже багато жовтка, в першій фазі гаструляціі двуслойний зародок утворюється шляхом розшарування, або деляминации,клітинного шару бластодерми. Цей тип освіти двуслойного зародка спостерігається у кишковопорожнинних, а також у вищих плацентарних ссавців, яйцеклітини яких втратили великі запаси жовтка. Гаструляція шляхом деляминации полягає в діленні клітин бластодерми, причому виникають при цьому дочірні клітини відтісняються в бластоцель і утворюють внутрішній шар клітин, або ентодерми, а зовнішній шар клітин, або колишня бластодерма, перетвориться в ектодерми. Коли відбувається відтискування дочірніх клітин в бластоцель, ми маємо справу з так званої первинної деляминации, а при вторинній деляминации клітини майбутньої ектодерми, навпаки, відтісняються кнаружи. При цьому способі гаструляціі бластопор не утворюється і гастроцель, природно, не повідомляється із зовнішнім середовищем.
Слід зазначити, що, незважаючи на своєрідність окремих способів гаструляції, в процесі утворення багатошарового зародка найчастіше спостерігається комбінація різних типів гаструляції - двох і навіть трьох типів.
Мезодерма, та способи її утворення.
Мезодерма — один з зародкових листків, які формуютьембріон тришарових тварин. Мезодерма формується в процесі гаструляції, завдяки міграції клітин з ектодерми, або внаслідок випинання та відокремлення карманів первинної кишки.
Еволюційно виникнення мезодерми було важливимароморфозом, що дав початок розвитку практично всіх тварин із складною будовою тіла. Виникнення мезодерми стало також і передумовою для розвитку целома (хоча він існує і не у всіх тришарових тварин).
Загалом, з мезодерми формуються такі системи органів:
Кістки
Більша частина кровоносної та лімфатичної систем, включаючи серце.
Сполучна тканина
Гладенькі м'язи кишкового тракту
Поперечносмугасті м'язи
Перитоніум
Репродуктивна система
Видільна система, включаючи нирки
Утворення третього шару клітин, або третього зародкового листка - мезодерми, в процесі ембріонального розвитку тварин здійснюється чотирма способами: телобластіческім, ентероцельним, ектодермального і змішаним, або перехідним.
У первичноротих тварин освіту мезодерми здійснюється за рахунок декількох великих клітин, розташованих під час гаструляції на кордоні між ектодерми і ентодерми з боків бластопор. Походження цих клітин не з'ясовано, так само як і ставлення їх до якогось із зародкових листків, оскільки вони відокремлюються вже на стадії дроблення. Дочірні клітини зазначених клітин розташовуються між ектодерми і ентодерми і формують третій шар клітин - мезодерми. Оскільки вихідні клітини, що утворюють мезодерму, протягом всього часу розташовуються на задньому кінці зародка, то відповідно до своєї локалізації вони отримали назвутелобластов, а сам спосіб утворення мезодерми - телобластіческого. Вторинна порожнина тіла зародка, або цілому, утворюється клітинами мезодерми внаслідок їх розшарування.
У Вториннороті, включаючи нижчих хордових, мезодерма утворюється з клітин ентодерми. При цьому способі утворення мезодерми клітини стінки первинної кишки, або ентодерми, інтенсивно розмножуючись, утворюють з боків первинної кишки мішкоподібні випинання в бластоцель. Ці випини, врости в бластоцель, розташовуються між ектодерми і ентодерми. Надалі ентодермальні випинання отшнуровиваются від ентодерми, а цілісність стінки первинної кишки і ентодермальних виростів відновлюється шляхом розмноження клітин. Після відокремлення від первинної кишки клітинний матеріал ентодермального походження іменується третім зародковим листком або мезодермою. Розростаючись, мезодерма заповнює всю порожнину бластоцелем, а порожнину мешкоподобних виростів, будучи за походженням порожниною гастроцель, іменується вторинної порожниною тіла. Цей спосіб утворення мезодерми отримав назвуентероцельного, оскільки мезодерма відбувається з ентодерми при одночасному утворенні целома.
У плазунів, птахів, ссавців тварин і людини третій зародковий листок утворюється з ектодерми під час другої фази гаструляції. Під час другої фази гаструляції спостерігається впячивание клітин ектодерми в бластоцель при одночасній імміграції клітин ектодерми в простір між ектодерми і ентодерми. Клітини, виселяють з ектодерми і розміщуються між останньою і шаром клітин ентодерми, є клітинами мезодерми. Цей спосіб утворення мезодерми називається ектодермального.
У хрящекостних риб та земноводних спостерігається змішаний ний, або перехідний, спосіб утворення мезодерми, який є відображенням еволюційних перетворень ембріонального розвитку від нижчих хордових до вищих. У цих тварин мезодерма формується в процесі гаструляції одночасно з ектодерми і ентодерми.
Пізня гаструла. Гісто- та органогенез.
На стадії пізньої гаструли зародок утворений двома шарами клітин - зовнішнім (первинної ектодерми) і внутрішнім (первинної ентодермою). У всіх тварин, крім двошарових (губки і кишковопорожнинні), утворюється третій шар - мезодерма, який у первичноротих відбувається з телобластов; біля Вториннороті матеріал мезодерми входить до складу первинних енто-або ектодерми і починає виокремлювати з них у період гаструляції. Відкриття стадії двошарового зародка в розвитку багатоклітинних мало важливе значення для доказу єдності походження тварин. Гістогене́з (від грец. histos — тканина + грец. genesis — утворення, розвиток) — сукупність процесів, що приводять до утворення і відновлення тканин в процесі індивідуального розвитку (онтогенезу).
В утворенні певного виду тканин бере участь той чи інший зародковий листок. Наприклад, м'язова тканина розвивається змезодерми, нервова — з ектодерми, і т. д. У ряді випадків тканини одного типу можуть мати різне походження, наприклад,епітелій шкіри має ектодермальне, а всмоктуючий кишковий епітелій — ентодермальне походження.
Гістогенез (від греч.(грецький) histos — тканина і ...генез ), розвиток тканин, сукупність закономірно протікаючих процесів, що забезпечують виникнення, існування і відновлення тканин тваринних організмів з їх специфічними в різних органах властивостями. Вивчення Р. різних тканин і його закономірностей — одне з найважливіших завдань гістології . Терміном «Г.» прийнято позначати розвиток тканин в онтогенезі . Проте закономірності Р. не можуть розглядатися у відриві від еволюційного розвитку тканин (філогистогенеза). У основі Р. лежить що починається з найраніших стадій ембріогенезуклітинна диференціювання — розвиток наростаючих морфо-функціональніх відмінностей між клітками, що спеціалізуються. Це складний молекулярно-генетичний процес закономірного включення активності генів, що визначають специфіку білкових синтезів в клітці. Розмноження кліток, їх взаїмоперемещенія і ін. процеси приводять до формування ембріональних зачатків, що є групами кліток, закономірно розташованими в телі зародка. В результаті тканинного диференціювання ембріональних зачатків виникає все різноманіття тканин різних органів тіла. У послезародишевом періоді процеси Р. підрозділяють на 3 основних типа: у тканинах, клітки яких не розмножуються (наприклад, нервова тканина); у тканинах, розмноження кліток яких пов'язане головним чином із зростанням органу (наприклад, паренхіма травних. залоз, нирок); у тканинах, що характеризуються постійним оновленням кліток (наприклад, кровотворна тканина багато покривних епітеліїв). Сукупність кліток, що здійснюють певний Р., підрозділяють на ряд послідовних груп (фондів): фонд родоначальних кліток, здібних як до диференціювання, так і до заповнення убули собі подібних; фонд кліток-попередників, що диференціюються і здібних до розмноження; фонд зрілих, таких, що закінчили диференціювання кліток. Відновлення пошкоджених або частково втрачених тканин після травм здійснюється завдяки т.з. репаратівному Р. За патологічних умов процеси Р. можуть піддатися глибоким якісним змінам і привести до розвитку пухлинних тканин
Органогене́з (від грец. органон - орган) — процеси утворення та розвитку органів у тварин. Розрізняють онтогенетичний органогенез, який досліджується ембріологією та біологією розвитку, і філогенетичний органогенез, що досліджується порівняльною анатомією. Окрім опису та аналізу протікання процесів органогенезу, до завдань вказаних дисциплін входять розкриття та пояснення причин цих процесів у філогенезі та онтогенезі. Порівняльна анатомія розглядає виникнення нових органів, їх перетворення, поділ, прогресивний розвиток та редукцію, процеси рудиментації та ін. Вивчення розвитку форми органів у зв'язку з їх функцією призвело до відкриття основних закономірностей філогенетичного органогенезу. Такими є принципи диференціації та інтеграції, а також зміни функцій як керівного початку в філогенетичній перебудові органів. Онтогенетиний органогенез до певної міри повторює філогенетичний органогенез (див. Біогенетичний закон). Під час першого здійснюється послідовне диференціювання та інтеграція органів, а також неравномірний ріст й активне переміщення клітинного матеріалу. Дослідження причин онтогенетичного органогенезу доступно точному вивченню, особливо завдяки можливості застосування експериментального методу (див. Експериментальна ембріологія). У рослин терміном «органогенез» звичайно позначають формування і розвиток основних органів (кореня, стебла, листь, квіток) у процесі онтогенезу з ділянки недиференційованої тканини — мерістеми.
Явище ембріональної індукції.
явище ембріональної індукції – коли одна з частин зародка (індуктор) спонукає напрям розвитку іншої. Ембріональний розвиток складається з наступних основних етапів: 1) дроблення, в результаті якого утворюється багатоклітинний зародок; 2) гаструляції, в процесі якої виникають перші ембріональні тканини — ектодерма і ентодерма, а зародок стає двошаровим; 3) первинного органогенезу — утворення комплексу осьових органів зародка — нервової трубки, хорди, кишкової трубки.
Ембріональну індукцію можна визначити як явище, при якому в процесі ембріогенеза один зачаток впливає на інший,визначаючи шлях його розвитку, і, крім того, сам піддається індуцируючому впливу з боку першого зачатка.
Сенситивні періоди ембріогенезу.
Адаптивний характер розвитку організму визначає необхідність врахування у віковій періодизації не тільки особливості морфо-функціонального розвитку фізіологічних систем організму, але і їх специфічної чутливості до різних зовнішніх впливів. Фізіологічними та психологічними дослідженнями показано, що чутливість до зовнішніх впливів носить вибірковий характер на різних етапах онтогенезу. Це стало основою уявлення про сенситивні періоди як періоди найбільшої чутливості до впливу факторів зовнішнього середовища, навчальних і фізичних навантажень. Своєрідним підтвердженням дії онтогенетичної програми розвитку потрібно вважати існування періодів підвищеної сприйнятливості організму до зовнішніх впливів. Саме в ці («критичні», чи «сенситивні») періоди найтісніше взаємодіють соціальні й біологічні програми індивідуального розвитку.
Поняття «сенситивні періоди» ввів у науку Г. Фріз, розуміючи під цим такий проміжок онтогенетичного розвитку, коли різні види живих істот виявляють високу чутливість до певних впливів середовища. Як приклад він наводить бджолу, що може перетворитися у матку лише при умові споживання певної їжі у певному віці; якщо ж та сама їжа буде дана пізніше цього періоду, то бджола маткою стати не може. На цьому ж наголошував і П. Морган, спостерігаючи за життям одомашнених снігурів. «Молодих снігурів, писав він, яких хочуть навчити співові особливо витончених мелодій, виймають з гнізда, коли у них починають рости пір'їнки на хвості, і годують лише насінням ріпи, розмоченим у воді і змішаним з білим хлібом».
Різні дослідження норми і патології людського розвитку, починаючи з ембріогенезу, дають підставу робити висновок про наявність сенситивних періодів і в людини. П. Г. Свєтлов на матеріалах ембріології з погляду критичних періодів розвитку з'ясовує принципи дії середовища на онтогенез. Головною ознакою критичних періодів розвитку, на його думку, є підвищення чутливості до місцевих агентів: «Універсальне поширення цього явища і його постійність наводить на думку, що природна періодичність, зумовлювана критичними періодами розвитку, повинна мати серйозне значення в онтогенезі».
Протягом вагітності за нормальних умов розрізняють такі критичні періоди. Перший період — це запліднення й етап імплантації (перший тиждень після запліднення), другий — ембріональний період, тобто збігається з часом, який передує утворенню плаценти. У цей час зародок продовжує живитись речовинами, що надходять із стінки матки, причому часто між ембріоном і матір'ю відбувається обмін несумісними білковими фракціями. Третій період збігається з бурхливим розвитком органів у першій половині плодового періоду. З'являється велика потреба плода в поживних речовинах і особливо в кисні. Кров матері насичується надмірною кількістю продуктів обміну, внаслідок чого можлива гіпоксія і токсинемія вагітної.
Плацента (placenta) є частиною плодових оболонок. Розвивається в ділянці трофобласта, який безпосередньо прилягає до стінки матки, протягом двох — п'яти місяців вагітності. Це орган, який здійснює процеси обміну між кров'ю матері й плода. У розвитку і формуванні плаценти бере участь трофобласт бластоцисти, який є основою утворення ворсинчастого хоріона, і слизова оболонка матки у вигляді основної відпадної (децидуальної) оболонки. При цьому замкнута капілярна мережа ворсин хоріона ніби плаває в крові лакунарних розширень цієї оболонки матки.
Плацента гальмує перехід деяких токсичних речовин із крові матері в кров плода. Разом з тим через плаценту в кров плода проникають алкоголь, наркотичні речовини, нікотин, гемолітичні отрути, ртуть, миш'як тощо. Бар'єрну функцію плацента має лише у фізіологічних умовах. Отруєння алкоголем, нікотином та іншими речовинами різко зменшує цю функцію, що призводить до захворювань, формування вад плода і навіть до його загибелі.
Особливості ембріонального розвитку людини.
Ембріогенез також Зародковий розвиток, Ембріональний розвиток,— розвиток організму тварини, що відбувається в оболонках яйця поза материнським організмом або усередині нього. Зародковому розвитку передує період передзародкового розвитку, коли зростає, формується і дозріває яйцеклітина. Після зародкового розвитку період післязародкового (постембріонального) розвитку. В ході зародкового розвитку з однієї відносно просто організованої яйцеклітини утворюється багатоклітинний організм, що складається з різних органів і тканин і здатний до самостійного існування. У деяких тварин, наприклад голкошкірих, зародки виходять з оболонок на дуже ранніх стадіях, і основні процеси розвитку проходять у них в постембріональний період. У всіх тварин зародковий розвиток складається з запліднення (або активації яйця при партеногенезі), дроблення, бластуляції, гаструляції, органогенезу і виходу з оболонок або народження. Залежно від біології розмноження (кількість яєць, тип запліднення, тривалість зародкового розвитку, джерела живлення зародка, міри турботи про потомство) будова яйця і характер зародкового розвитку варіює.
У процесі ембріонального розвитку людини зберігаються загальні закономірності розвитку і стадії, характерні для хребетних тварин. Разом з тим з'являються особливості, що відрізняють розвиток людини від розвитку інших представників хребетних; знання цих особливостей необхідно лікарю. Процес внутрішньоутробного розвитку зародка людини продовжується в середньому 280 доби (10 місячних місяців). Ембріональний розвиток людини можна розділити на три періоди: початковий (1-я тиждень розвитку), зародковий (2-8-я тиждень розвитку), плодовий (з 9-го тижня розвитку до народження дитини). До кінця зародкового періоду закінчується закладка основних ембріональних зачатків тканин і органів і зародок здобуває основні риси, характерні для людини. До 9-й тижня розвитку (початок 3-го місяця) довжина зародка складає 40 мм, а маса близько 5 р. У курсі ембріології людини, досліджуваній на кафедрі гістології та ембріології, основна увага приділяється особливостям статевих кліток людини, запліднення і розвитку людини на ранніх стадіях (початковий і зародковий періоди), коли відбуваються утворення зиготи, дроблення, гаструляція, формування зачатків осьових органів і зародкових оболонок, гістогенез і органогенез, а також взаємодії в системі мати - плід.
Провізорні органи, особливості структури та функціональна роль.
Провізорні органи (нім. provisorisch — попередній, тимчасовий, від лат.provideo — передбачаю, наперед турбуюсь) — тимчасові органи у зародків та личинок тваринних організмів, які зникають у процесі їхнього розвитку. Одна з форм ценогенезу, або ембріоадаптації.
Провізорні органи, або тимчасові, розвиваються в процесі ембріогенезу поза тілом зародка, виконують численні функції, що забезпечують розвиток і ріст самого зародка. До провізорних органів птахів відносяться: жовточний мішок, амніон, серозна оболонка, алантоїс. Провізорні органи забезпечують найважливіші функції організму, що розвивається, до формування та початку функціонування дефінітивних органів, характерних для дорослих особин. Провізорні органи сучасних організмів у ряді випадків дозволяють передбачати організацію предкових форм, що допомагає встановлювати еволюцію певних груп тварин.
Прикладами провізорних органів є:
черевні кінцівки та зябра личинок комах;
зябра, ротові придатки та хвіст пуголовків земноводних;
судини жовточного мішка у зародків риб;
амніон, алантоїс та серозна оболонка у амніот.
Амніон. Жовточний мішок Алантоїс.
Амніо́н — одна із зародкових оболонок у амніот (плазунів, птахів, ссавців).
Амніон — тонка напівпрозора мембрана, яка оточує плід з усіх боків і створює навколишнє середовище плода протягом його внутрішньо-маткового життя. Внутрішня стінка — епітелій, який утворився з ектодерми а зовнішня — сполучнотканинна основа, сформована з мезодерми. Амніон заповнений прозорою водянистою рідиною, яка містить солі NaCI, КСІ, білки, незамінні амінокислоти (аланін, лейцин, гістідін, аргінін лізин), вуглеводи, жири, ферменти, гормони, простагландини, гістамін серотонін, катехоламін, деякі фактори росту (пантотенова і фолієва кислоти).
Спочатку вагітності амніотична рідина є трансудатом трофобласта, потім її секретує амніотичний епітелій, легені, шлунково-кишковий тракт плода, а у телят — епітеліальні бляшки, розмішені навколо пупка. Вона постійно відновлюється, 3-4 рази на добу виробляється і поглинається. У першу половину вагітності кількість амніотичної рідини збільшується досягаючи на 5-6 міс. у великих тварин 6-7 кг, а в другу — зменшується. тому, що плід п'є її для підтримки свого водного балансу. На початку вагітності амніотична рідина має світлий колір, пізніше вона біліє, оскільки у ній з'являються жирові частинки. Може бути коричневатою при передчасному виході меконію або червонуватою після смерті чи розпаду плода.
Розвивається амніон із зовнішніх і середедніх зародкових листків.
У птахів і ссавців в амніоні є кровоносні судини і скоротливі м'язові елементи. Порожнина амніона наповнена амніотичною рідиною, або плодовою водою, що захищає зародок від висихання, механічних ушкоджень, дозволяє йому вільно рухатись, бере участь в обміні речовин плоду. Амніон є також у деяких комах.
Жовточний мішок утворюється в результаті розростання клітин гіпобласта (позазародкова ентодерма) у ширину з поступовим обростанням усього жовтка. Після виникнення хордомезодермального зачатка між екто — і ентодермою проростають парієтальний і вісцеральний листки мезодерми. Зародок піднімається над жовтком і відмежовується від нього за допомогою тулубової складки. При утворенні тулубової складки зародкова ентодерма, що була до того розпластана на жовтку, скручується в кишкову трубку.
Зародок з’єднується з жовточним мішком за допомогою порожнистого канатика — жовточної стеблинки. Жовточний мішок виконує трофічну функцію і кровотворну (у ньому утворюються перші клітини крові), а також перші судини і первинні статеві клітини — гонобласти.
АЛАНТОЇС (грец. allantoeides — ковбасоподібний) — ковбасоподібний зародковий орган вищих хребетних. Зачаток А. присутній у амфібій. У зародків рептилій, птахів та ссавців закладка А. являє собою випинання заднього відділу ентодермальної кишки, що надалі виростає далеко за межі тіла самого зародка і стає важливим органом зв’язку організму, що росте, з середовищем. А. складається з двох клітинних шарів — ентодерми та кишкового листка мезодерми. У рептилій та птахів А. розташовується під поверхнею яйцевої шкаралупи. Дрібнопетляста мережа кровоносних судин, що лежить на його поверхні, через серозну оболонку входить у тісний контакт із внутрішньою поверхнею дрібнопористої шкаралупи, чим забезпечується газообмін з атмосферою. Порожнина А. у рептилій, птахів та багатьох ссавців зв’язана з порожниною кишечнику каналом, що відкривається в клоаку, тому в порожнину А. надходять продукти виділення. Початковий відділ А. далі утворює сечовий міхур. У вищих ссавців А. у середині зародкового періоду стає органом дихання. У сумчастих ссавців судинна система, що бере участь у живленні та диханні зародка, належить цілком жовточному мішку, у плацентарних — головного значення набуває судинна система А.; судинне поле жовточного мішка може мати лише допоміжне значення. У плацентарних, які мають так звану плаценту, що не відпадає, А. добре розвинутий. У вищих ссавців, що мають плаценту, яка відпадає, ентодерма А. більш-менш скорочена, кровоносні судини добре розвинуті. У людського зародка ентодермальна закладка А. вже на ранніх стадіях різко скорочена. У навколишньої закладки А. зародкової сполучної тканини, що формує основу так званої черевної ніжки, добре розвиваються кровоносні судини А. (надалі — пупкові вена й артерії). Черевна ніжка є закладкою пупкового канатика. Вважається, що редукція А. у людини є наслідком інтерстиціальної імплантації, що настає раніше: вже наприкінці 6-ї доби після запліднення людський зародок, що має вид малесенького сочевицеподібного тільця (близько ¼ мм у діаметрі), у слизовій оболонці матки та у її товщі проходить наступні стадії розвитку. Основа виросту А. у пізнього людського зародка, як і у всіх ссавців, перетворюється на сечовий міхур, а канал А., що йде до пупкового отвору, у так званий первинний сечовід, чи urachus, зберігається у дорослої людини у вигляді середньої пупково-пухирчастої зв’язки (ligamentum vesico-umbilicale medium).
Плацента, її утворення та значення
Плацента (лат. placenta, від греч.(грецький) Plakús — коржик), дитяче місце, у людини, майже у всіх ссавців, а також у деяких хордових і безхребетних тварин — орган, що здійснює зв'язок і обмін речовин між організмом матері і зародком в період внутріутробного розвитку (див. Живонародження ) . Через П. зародок отримує кисень, а також живильні речовини з крові матері, виділяючи в неї продукти розпаду і двоокис вуглецю. П. виконує і бар'єрну функцію, активно регулюючи вступ різних речовин в зародок. У П. містяться ферменти, що беруть участь в обміні речовин зародка, вітаміни.
Плацентація — це утворення плодових оболонок.
Трофобласт росте дуже швидко, а ембріобласт під своїм тягарем опускається всередину, і над ним з'являється склепіння складок трофобласта. Поступово це склепіння стуляється, і між складками, що зімкнулися, залишається лише невеличкий отвір — пупок амніона. 3 часом і він заростає, внаслідок чого ембріон, опинившись у центрі новоутвореної порожнини, стає ізольованим від зовнішнього середовища ділянки поверхні трофобласта, які зімкнулися, атрофуються, і таким чином з трофобласта утворюється зовнішня оболонка — трофобласт і внутрішня, яка повністю оточує ембріон і в ділянці пупка переходить у шкіру — водна оболонка, або амніон (від грец amnio чаша для жертовної крові) 18 денний зародок великої рогатої худоби оточений амніоном. Весь зародковий пухирець (трофобласт) швидко витягується в довжину У 16 днів він займає 2/3 довжини рога матки (від 70 до 240 мм), на 17-и день — повністю вагітний ріг, на 19-20-и день — досягає верхівки протилежного рога матки його товщина становить близько 2 мм. На поверхні трофобласта на 20-й день утворюються ворсинки і з цього часу він стає прохоріоном. Коли до прохоріона підходять судини, що виходять з черевної порожнини плода разом з сечовою оболонкою і в ворсинки прохоріона приблизно на 30-й день вростають капіляри, прохоріон називається хоріоном (грен, chonon — послід) або судинною оболонкою. Дрібні кровоносні судини збираються від всіх ворсинок разом і через пуповину ведуть до плода
Простір між амніоном і хоріоном заповнюється магмою з первинної кишки утворюється алантоїс (грец allantois — ковбасовидний), або сечова оболонка. Алантоїс бере початок від верхівки сечового міхура і проходить через пупковий отвір у вигляді сечової протоки, або урахуса. Він збільшується у міру наповнення первинною сечею, розповсюджується між водною і судинною оболонками і до 26-го дня огортає ембріон майже з усіх боків
Взаємовідношення плодових оболонок у м'ясоїдних, кролиць і кобил алантоїс займає весь простір між хоріоном і амніоном. Зовнішній листок, зливаючись з хоріоном, формує аланто -хоріон, тоді як внутрішній листок, більше чи менше примикаючи до зовнішньої поверхні амніона, утворює аланто – амніон. Тільки на невеликій площі з вентральної поверхні алантоїс не покриває амніон у цьому місці виходить пупковий канатик. Плід, замкнений в аланто-амніон, може вільно пересуватися разом з останнім у порожнині сечової оболонки. Це сприяє народженню плода у плодовій оболонці без її розриву (у сорочці).
У свиней і жуйних алантоїс оточує амніон двома рогами з боку черева плода і не покриває спинку плода.
При багатоплідній вагітності у всіх тварин кожен плід має зовсім відособлені водну, сечову і часто навіть судинну оболонки. У корів при багатоплідній вагітності судинна оболонка часто буває загальною, але з відокремленими для кожного плода зонами розповсюдження кровоносних судин і плацентарних зв'язків. При утворенні анастомозів між судинами різних плодів кров одного плода вільно попада? до кровоносної системи другого, і у випадку різностатевої двійні гормони бичка затримують розвиток статевих органів у телички. Останні народжуються з недорозвиненими статевими органами фримартинами і остаються неплідними. Це пов'язано з порівняно швидшим формуванням в ембріональний період зародка чоловічої статі.
У овець і кіз при багатоплідній вагітності загальна судинна оболонка зустрічається нерідко, але анастомози не утворюються, і тому фримартинізм майже не спостерігається.
У свиней і м'ясоїдних судинні оболонки завжди відокремлені.
Види плацент. їх морфологічна особливість.
Класифікація плацент
1. За розміщенням ворсинок (за типом плаценти):
— розсіяна, або дифузна pi. diffusa s. pi. disseminate (кобила, свиня);
— множинна pi. multiplex (жуйні);
— зональна, або пояскоподібна pi. zonaria має вигляд пояска шириною 2-3 см (м'ясоїдні);
— дископодібна pi. discoidea (примати, кролиці, гризуни).
2. За характером зв'язку плодової і материнської частин плаценти:
— ахоріальна (кенгуру, кити);
— епітеліохоріальна (кобила, свиня);
— десмохоріальна, тобто сполучнотканинна (жуйні);
— ендотеліохоріальна (м'ясоїдні);
— гемохоріальна (примати).
3. За типом відокремлення плацент у період пологів:
— невідпадаюча pi. adecidua — при її відділенні слизова оболонка матки не пошкоджується і пологи проходять майже без кровотечі. Вона характерна для тварин з епітеліохоріальною і десмохоріальною плацентою;
— відпадаюча pi. decidua — характерна для приматів, кролиці, гризунів. У них пологи проходять з розривами слизової оболонки матки і з досить виразною кровотечею.
4. За характером живлення плода
— гістіотрофна, властива для приматів і м'ясоїдних, які живляться тканинними соками (гістіотрофом),
— ембріотрофна, у жуйних, свиней, кобил (живлення матковим молочком).
Загальні уявлення про тканини організму.
Ткани́на — сукупність клітин, не обов'язково ідентичних, але спільного походження, що разом виконують спільну функцію. Тканинний рівень — це рівень клітинноїорганізації, проміжний відносно клітин та усього організму. Органи утворюються функціональним об'єднанням тканин одного чи кількох видів.
Вивченням тканин займається наука гістологія, або ж, у зв'язку із захворюваннями,гістопатологія. До набору класичних матеріалів та інструментів для дослідження тканин входять парафіновий блок, у який вводиться тканина для фіксації і зручності одержання зрізу, гістологічний барвник та оптичний мікроскоп. За останні десятиліття розвитокелектронної мікроскопії, іммунофлюоресценції та використання заморожених зрізів тканин уможливив детальніше візуальне дослідження тканин. Ці інструменти дозволяють спостерігати звичайний вигляд здорових і уражених тканин, цим самим суттєво покращуючи клінічне діагностування та прогнозування. Ці досягнення використовує сучасна медицина, зокрема у хірургії — гістологічне дослідження є базовим для тканинної діагностики пухлин, інших паталогічних станів.
В організмі тварин та людини розрізняють 4 типи тканин:
сполучна тканина
м'язова тканина
нервова тканина
епітеліальна тканина.
У багатоклітинному організмі людини є клітини, які відрізняються за своєю будовою та функціями, що зумовлено їхньоюдиференціацією (з латин. різний, відмінний) та спеціалізацією при виконанні певних функцій. Диференціація та спеціалізація клітин генетично запрограмовані. Змінити це неможливо. Так, нервова клітина ніколи не виконуватиме функцію еритроцита, а клітини, що виробляють травні соки, не стануть кістковими чи м’язовими. Окремi групи клітин утворюють певну тканину.
У людини розрiзняють чотири типи тканин: епiтелiальну, нервову, м’язову і сполучну.
Епітеліальна тканина (з грец. над та сосок) утворює зовнiшнi покриви тiла — елементи шкiри; слизові оболонки дихальних i травних шляхiв, а також внутрішні оболонки серця й судин; із неї побудованi легеневі міхурці (альвеоли), залози внутрiшньої секрецiї, а також шкірні (потові, сальні), молочні, слізні, статеві залози.
Епiтелiальна тканина захищає iншi тканини, що мiстяться пiд нею. Зовнiшнi клiтини епiтелiю шкіри роговiють і відмирають.
Наприклад, коли людина багато пише, то на середньому пальцi її правої (а в шульгiв — лiвої) руки утворюється мозоля.
Крiм того, епiтелiальна тканина виконує функцiї видiлення й всмоктування (епiтелiй шлунка, кишечнику). Вона має високу здатнiсть до вiдновлення.
Епітеліальна тканина — не тільки покривна, а й погранична. Цим зумовлена її участь в обмінних процесах: газообмін через епітелій альвеол легенів; усмоктування поживних речовин у тонкому кишечнику.
Є епітелій багатошаровий (роговіючий, нероговіючий і перехідний) та одношаровий (циліндричний, кубічний, плоский). Шкіра вкрита роговіючим багатошаровим плоским епітелієм, а слизова оболонка шлунка та дихальних шляхів — одношаровим циліндричним.
Сполучна тканина є насамперед опорною, оскільки вона бере участь в утвореннi кiсток, хрящiв, зв’язок, мiжклiтинної сполучної речовини, пiдшкiрної основи, сухожиль, зубiв. Вона здебiльшого не потребує багато кисню та поживних речовин, тому мiстить незначну кiлькiсть кровоносних судин, а процеси обмiну речовин у нiй відбуваються досить повiльно.
Сполучна тканина дуже різниться за будовою та функціями. Для неї характерна наявність клітин і міжклітинної речовини; остання складається з волокон та основної речовини. Розрізняють кілька видів сполучної тканини.
Пухка волокниста сполучна тканина складається з клітин та безладно розміщених в основній речовині волокон. Вона переважно знаходиться вздовж кровоносних судин. Її різновидом є ретикулярна сполучна тканина. Пухка волокниста сполучна тканина утворює основу кровотворних органів та органів імунної системи (кістковий мозок, селезінка, лімфовузли).
Щільна волокниста сполучна тканина має небагато клітин, які розміщені між численними сполучнотканинними волокнами, що густо переплітаються. Із неї побудовані зв’язки та сухожилля.
Хрящова тканина складається з хрящових клітин (хондроцитів), які розміщуються по 2–3 серед основної речовини, що має консистенцію надзвичайно щільного гелю.
Кісткова тканина відрізняється особливою щільністю та особливими механічними властивостями; вона складається з кісткових клітин, замурованих у звапнену міжклітинну речовину.
Окремо треба вiдзначити роль жирової сполучної тканини, що становить пiдшкiрну основу, бо це — своєрiдне енергетичне депо організму. Крiм того, вона захищає внутрiшнi органи вiд механiчних пошкоджень. Як добрий теплоiзолятор, жировий прошарок сприяє збереженню тепла в органiзмi.
Своєрiдний вид сполучної тканини — кров, основна речовина якої — плазма — має рiдку консистенцiю. У нiй вiльно плавають клiтиннi елементи.
Вивченню ролi сполучної тканини в органiзмi присвятив багато наукових праць видатний український фiзiолог Олександр Богомолець.
М’язова тканина — основний елемент м’язiв; вона забезпечує процеси руху. В органiзмi людини 40 % його маси становлять м’язи. М’язова тканина має спеціальні скоротливі волокна — міофібрили. Розрiзняють посмуговану (поперечносмугасту), непосмуговану(гладеньку) i серцеву м’язові тканини.
Із посмугованої м’язової тканини складаються скелетнi м’язи, які скорочуються довільно (свідомо) під впливом нервових імпульсів, що надходять із головного мозку. М’язова тканина складається з багатоядерних клітин, що утворюють посмуговані м’язові волокна складної будови. Через особливе розташування міофібрил у цитоплазмі цих клітин, під мікроскопом видно чергування світлих і темних ділянок уздовж м’язового волокна. Скелетні м’язи забезпечують пересування людини, а також рухи частин її тіла.
Непосмуговані м’язи складаються з клітин веретеноподібної форми, які утворюють середню моторну (з латин. рухливий) стiнку шлунка, кишечнику, матки, кровоносних судин. Вони виконують функцiю проштовхування, наприклад, їжi в травних шляхах, або сечi в сечових, чи плоду маткою, коли настає час народитися дитині. Цi м’язи не пiдкоряються нашiй волi.
Серцевий м’яз подібний до посмугованих і непосмугованих м’язів. За будовою він подібний на скелетний м’яз, але виконує функції непосмугованого — забезпечує рух крові по судинах. Людина не може керувати роботою серцевого м’яза. Для нього характерне функціональне з’єднання розташованих
поряд клітин за допомогою спеціальних контактів. Така його будова забезпечує скорочення однойменних відділів серця (передсердь, шлуночків) як єдиного цілого. У клітинах серцевого м’яза є велика кількість мітохондрій.
Нервова тканина — основна тканина центральної та периферичної нервової системи — має надзвичайно складну будову. Це приблизно 10–14 млрд нервових клiтин (нейронiв), тіла яких утворюють сiру речовину головного та спинного мозку. Вони мають переважно зiрчасту або веретеноподiбну форму з вiдростками.
Здебільшого короткi вiдростки нервових клітин — дендрити (з грец. дерево) — сприймають і передають iнформацiю до тіла своєї клiтини, а довгi — аксони (з латин. вісь) — передають нервові імпульси до інших нейронів, м’язiв і секреторних клітин.
Інша частина нервової тканини — це нейроглія. Вона складається з клітин, що оточують нейрони. Нейроглія виконує опорну функцію та функцію живлення (трофічну) для нейронів. Кількість клітин нейроглії приблизно в 10 разів перевищує кількість нейронів. Відростки нервових клітин і клітини нейроглії утворюють білу речовину головного й спинного мозку.
Основними властивостями нервової тканини є збудливість і провідність.
Поняття про диферениіацію клітин.
Диференціація – це стійке структурно-функціональне перетворення клітин у різні спеціалізовані клітини. Диференціація клітин біохімічно пов’язана з синтезом специфічних білків, а цитологічних – з утворенням спеціальних органел та включень. При диференціюванні клітин відбувається виборча активація генів. Важливим показником клітинної диференціювання є зрушення ядерно-цитоплазмових відносини в бік переважання розмірів цитоплазми над розміром ядра. Диференціація відбувається на всіх етапах онтогенезу. Особливо чітко виражені процеси диференціації клітин на етапі розвитку тканин з матеріалу ембріональних зачатків. Спеціалізація клітин обумовлена їх детермінацій.
Детермінація – це процес визначення шляху, напрями, програми розвитку матеріалу ембріональних зачатків з утворенням спеціалізованих тканин. Детермінація може бути оотіпіческой (програмує розвиток з яйцеклітини і зиготи організму в цілому), зачаткових (програмує розвиток органів або систем, що виникають з ембріональних зачатків), тканинної (програмує розвиток даної спеціалізованої тканини) і клітинної (програмує диференціювання конкретних клітин). Розрізняють детермінацію: 1) лабільну, нестійку, оборотну і 2) стабільну, стійку і необоротну. При детермінації тканинних клітин відбувається стійке закріплення їх властивостей, внаслідок чого тканини втрачають здатність до взаємного перетворення (метаплазії). Механізм детермінації пов’язаний зі стійкими змінами процесів репресії (блокування) і експресії (деблокування) різних генів.
Клітинна загибель – широко поширене явище як в ембріогенезі, так і в ембріональному гістогенез. Як правило, у розвитку зародка і тканин загибель клітин протікає по типу апоптозу. Прикладами программированной загибелі є загибель епітеліоцитів в міжпальцевих проміжках, загибель клітин по краю зростаються піднебінних перегородок. Програмована загибель клітин хвоста відбувається при метаморфозі личинки жаби. Це приклади морфогенетичні загибелі. В ембріональному гістогенез також спостерігається загибель клітин, наприклад при розвитку нервової тканини, скелетної м’язової тканини та ін Це приклади гістогенетичної загибелі. В дефінітивного організмі шляхом апоптозу гинуть лімфоцити при їх селекції в тимусі, клітини оболонок фолікулів яєчників в процесі їх відбору для овуляції і ін
диференціювання клітин
Поняття про дифферона. У міру розвитку тканин з матеріалу ембріональних зачатків виникає клітинне співтовариство, в якому виділяються клітини різного ступеня зрілості. Сукупність клітинних форм, що становлять лінію диференціювання, називають дифферона, або гістогенетичному поруч. Дифферона становлять кілька груп клітин: 1) стовбурові клітини, 2) клітини-попередники, 3) зрілі диференційовані клітини, 4) старіючі і відмирають клітини. Стовбурові клітини – вихідні клітини гістогенетичному ряду – це самопідтримується популяція клітин, здатних диференціюватися в різних напрямках. Маючи високі проліферативними потенціями, самі вони (тим не менше) діляться дуже рідко.
Клітини-попередники (напівстовбурові, камбіальні) складають наступну частину гістогенетичному ряду. Ці клітини зазнають кілька циклів ділення, поповнюючи клітинну сукупність новими елементами, і частина з них потім починають специфічну диференціювання (під впливом факторів мікрооточення). Це популяція коммітірованних клітин, здатна диференціюватися в певному напрямку.
Зрілі функціонуючі і старіючі клітини завершують гистогенетический ряд, або дифферона. Співвідношення клітин різного ступеня зрілості в дифферона зрілих тканин організму неоднаково і залежить від основних закономірних процесів фізіологічної регенерації, властивих конкретному виду тканини. Так, в оновлюються тканинах виявляються всі частини клітинного дифферона – від стовбурової до високодиференційованою і гине. У типі зростаючих тканин переважають процеси росту. Одночасно в тканини присутні клітини середньої та кінцевої частин дифферона. В гістогенез мітотична активність клітин поступово знижується до низької або вкрай низькою, наявність стволових клітин мається на увазі тільки в складі ембріональних зачатків. Нащадки стовбурових клітин деякий час існують як проліферативний пул тканини, але їх популяція швидко витрачається в постнатальному онтогенезі. В стабільному типі тканин є лише клітини високодиференційованою і гине частин дифферона, стовбурові клітини виявляються лише в складі ембріональних зачатків і повністю витрачаються в ембріогенезі.
Вивчення тканин з позицій їх клітинно-дифферона складу дозволяє розрізняти монодіфферонние – (наприклад, хрящова, щільна оформлена сполучна тощо) і полідіфферонние (наприклад, епідерміс, кров, пухка волокниста сполучна, кісткова) тканини. Отже, незважаючи на те, що в ембріональному гістогенез тканини закладаються як монодіфферонние, надалі більшість дефінітивного тканин формуються як системи взаємодіючих клітин (клітинних дифферона), джерелом розвитку яких є стовбурові клітини різних ембріональних зачатків.
Тканина – це філо-і онтогенетично склалася система клітинних дифферона та їх неклітинних похідних, функції і Регенераторна здатність якої визначається гістогенетичному властивостями провідного клітинного дифферона.
Тканина є структурним компонентом органу і в той же час частиною однієї з чотирьох тканинних систем – покривних, тканин внутрішнього середовища, м’язових і невральних.
Види клітин: стовбурові, напівстовбуро, диференційовані.
Процес, який веде до утворення різних клітин з початково однорідних, у цитології називається диференціацією клітин. Процес диференціації клітин здійснюється як у період ембріонального розвитку організму, так і під час його постнатального життя. В ембріональний період (ембріогенез) ці процеси відбуваються як на рівні ембріональних зачатків, так і під час спеціалізації клітин і тканин.
Розвиток організму супроводжується двома процесами: клітинною проліферацією (від лат. proles — нащадок, потомок і phero — несу) — розростанням тканин унаслідок багаторазового поділу клітин та диференціацією (від лат. differentia — відмінність) — утворенням різних клітин з початково однорідних.
Диференціація клітин (цитодиференціація) звичайно наступає після проліферації. Клітини, які швидко розвиваються є, як правило, малодиференційованими (наприклад, мезенхімні клітини або клітини базального шару епітелію шкіри). Високодиференційовані клітини звичайно втрачають здатність до проліферації. Проліферацію, як і ріст, стимулюють різні гуморальні фактори, зокрема фактори росту.
Цитодиференціація — поява відмінностей між клітинами у багатьох тварин є незворотним процесом. Вона здійснюється лише в одному напрямку — від менш диференційованої до більш диференційованої клітини. Результатом диференціації є спеціалізована клітина конкретної морфології, здатна виконувати певні, визначені функції. Цитодиференціація у рослин і нижчих тварин — процес зворотний.
а) теорії клітинної диференціації
Якщо розглядати процес диференціації з точки зору генної теорії, то необхідно відзначити, що диференційовані клітини містять такий же набір генів (генотип), як і недиференційовані. Проте, переважна більшість генів неактивні, заблоковані. У процесі диференціації, з одного боку, включаються гени, під дією яких клітина повинна перетворитися в певний один тип, а з другого боку — репресувати (пригнічувати) ті гени, які могли б спрямувати її по іншому шляху диференціації. При диференціації клітини експресують суворо визначену частину геному, транскрибують специфічні РНК і синтезують специфічні білки, що й визначає морфологічні та функціональні ознаки спеціалізації клітин. Отже, відмінності між клітинами, які мають однаковий набір генів, визначає диференціальна активність генів.
Диференціація клітин приводить до утворення ліній з різним ступенем диференціації. Серед мільйонів клітин організму вдається визначити під мікроскопом приблизно 100 різних типів. Диференційовані клітини характеризуються морфологічними і особливо функціональними властивостями, що обумовлено специфічними властивостями білків. Отже, в основі будь-якої диференціації лежать структурні зміни білків. Оскільки білковим синтезом керує ДНК, то тим самим вона управляє диференціацією. Однак, жорстка програма синтезу білка, визначена ДНК, модифікується низкою умов. Реалізацію дії структурних генів, які беруть участь у диференціації, здійснюють гени-регулятори, зокрема для диференціації необхідна узгоджена відрегульована дія активацій і депресій різних генів, гра, яка регулювала б послідовність потенціальної активності генів.
б) типи морфологічної диференціації
Диференціація клітин відбувається, починаючи з ранніх стадій ембріогенезу, і продовжується формуванням тканин. Під час диференціації наступають зміни в цитоплазмі клітин внаслідок її взаємодії з ядром. Проте, найбільш помітною є морфологічна диференціація.
У ході розвитку виділяють ряд етапів (типів) морфологічної диференціації:
1) Оотипічна, яка проявляється вже під час запліднення. Особливо наглядно вона виступає в зиготі амфібій. На анімальному полюсі появляється так званий сірий серп, а в базальному — нагромаджується жовток.
2) Бластомерна — на рівні перших декількох дроблень. У деяких тварин чітко помітна різниця між окремими групами бластомерів, у жаби — це мікробластомери і макробластомери.
3) Зачаткова. У період утворення мезодерми відзначається різниця між клітинами окремих зачатків, коли в сомітах вони інші, ніж у сегментних ніжках чи спланхнотомі.
4) Тканинний тип диференціації продовжується весь період ембріонального і навіть постнатального розвитку. Найбільш помітна така диференціація в процесі розвитку мезенхіми, з якої утворюються всі види сполучної тканини (пухка, щільна, хрящова, кісткова) і формені елементи крові (еритроцити, всі види лейкоцитів).
Детермінація. У ході диференціації клітини певної тканини реалізують закріплені детермінацією потенції. Детермінація клітин(тканин) (від лат. determinatio — визначення) — процес, який програмує напрямок розвитку клітин, характерний для даного виду тканин. У результаті детермінації компетентна клітинна система обирає один з багатьох можливих шляхів розвитку. Отже, детермінація клітини — це процес визначення шляху її дальшого розвитку, виникнення якісної своєрідності клітин.
Клітина, детермінована в одному напрямку, продукує клітини, детерміновані таким же чином: “подібне породжує подібне”. Немає єдиної теорії механізму детермінації. Вважають, що детермінація відбувається внаслідок дії низки факторів. Оскільки в процесі детермінації не має значення блокування генів, бо під час реплікації ДНК всі гени повинні бути деблоковані, то виникає думка, що детермінованість у різних клітинних лініях мабуть залежить від постійності цитоплазми протягом послідовних поколінь. Цитоплазма впливає певним чином на активацію або гальмування генів, що приводить до синтезу певних молекул РНК, а тим самим і синтезу білків відповідних даному типові клітин.
Потенції клітин. У процесі диференціації відбувається поступове обмеження (рестрикція) потенцій клітин, у результаті чого клітини набувають структурних і функціональних властивостей зрілих.
Клітини організму відрізняються своєю потенцією (від лат. potentia — сила), “можливістю, яка не обов’язково реалізована”.Тотипотентні (від лат. totus — весь), або омніпотентні (від грец. omnis — кожний) клітини — це запліднені яйцеклітини, які служать предковими клітинами для всіх інших клітин організму; тотипотентними є практично усі рослинні клітини. Поліпотентні (від грец. poli — багато), або плюрипотентні (від лат. pluralis — множинний), чи стовбурові клітини частково втратили свої потенції, але з них можуть утворюватися багато типів клітин, наприклад, мезенхімні клітини, які дають початок клітинам крові, власне сполучної тканини, хрящової, кісткової та гладкої м’язової тканини. Уніпотентні (від грец. unis — один) клітини не зберігають потенцій і лише здатні утворювати собі подібні клітини, наприклад, клітини епітелію шкіри.
Стовбурові клітини — найменш диференційовані клітини певної тканини, зі збереженими високими потенціями, які є джерелом розвитку інших клітин тканин. Вони наявні у всіх тканинах під час їх ембріонального розвитку і зберігаються в багатьох тканинах зрілих організмів. Стовбурові клітини мають ряд характерних особливостей. Вони: (1) утворюють клітинну популяцію, яка сама себе підтримує; (2) рідко діляться; (3) стійкі до дії шкідливих факторів; (4) у деяких тканинах вони плюрипотентні, тобто можуть стати джерелом розвитку декількох видів диференційованих клітин.
Диферон. У тканинах звичайно присутні клітини різного ступеня диференціації, серед них є стовбурові, родоначальні (напівстовбурові), диференційовані, старі. Таку закономірну сукупність клітин певного типу тканини називають дифероном. Диферон (від лат. differo — розповсюджувати), або гістогенетичний ряд — це угруповання всіх клітин, які складають ту чи іншу лінію диференціації — від найменш диференційованих (стовбурових) до найбільш зрілих — диференційованих.
Взаємодія клітин у тканинах та міжгкнинні взаємодії.
Для всіх клітин є характерною здатність до розмноження, регенерації, регуляції функцій, росту, реплікації молекул, функціональної спеціалізації. їм властива збудливість, провідність, активний транспорт речовин, секреція й екскреція, скоротливість.
Клітини поєднуються в тканини і злагоджено функціонують завдяки спеціальним взаємодіям. Це починається ще під час ембріогенезу на основі функціонування спеціальних генетичних програм, що забезпечують диференціювання клітин і наявність на їх поверхні рецепторів-маркерів. Завдяки їм клітини з’єднуються спеціальними контактами. Простий контакт — зближення плазмалем сусідніх клітин на відстань 10-20 нм, щільний контакт— мембрани в деяких місцях зближуються до 2-3 нм; десмосома — спеціальний контакт, до якого з боку цитоплазми контактуючих клітин підходять фибрилярні елементи цитоске- лета. Такі контакти забезпечують велику силу зчеплення клітин. Не лише механічне з’єднання, але й функціональний зв’язок між клітинами забезпечують: 1) нексус (щілинне з’єднання), коли рлазматичні мембрани розділені проміжком 2-3 нм; ділянка контакту має канали обміну невеликими молекулами, утому числі і сигнальними; 2) синапс — характерний для нервової тканини. У спеціальних ділянках контакту між нейронами утворюються нервово-м’язові і нервово-епітеліальні контакти. Сигнальними молекулами тут є спеціальні хімічні речовини — адреналін, но- радреналін, ацетилхолін, що діють на рецептори і спричинюють функціональну відповідь клітини.
Між клітинами діє також система сигналізації:
Віддалена, через рідину внутрішнього середовища (наприклад, ендокринна).
Контактна, що діє між близько розташованими клітинами за допомогою спеціальних сигнальних молекул (локальних хімічних медіаторів, що руйнуються швидко, одразу після того, як встигли вплинути на клітини ближнього оточення).
Контактна через щілинні з’єднання сусідніх клітин. (наприклад, в серцевому м’язі всі кардіоміоцити мають тісний фізіологічний контакт.
Зовнішнім сигналом для клітини є хімічні речовини. Реагують на хімічні сигнали лише ті клітини, які мають спеціальні рецептори. Відомо два різновиди клітинних рецепторів, розташованих на поверхні цитоплазматичних мембран, а саме:
Рецептори-канали. Це чуттєві до гормонів та нейромедіаторів білки, що утворюють іонні канали.
Рецептори-ферменти. Білки, які шляхом фосфорилювання активують білки клітини у відповідь на зв’язування сигнальної молекули.
Тканини — система однорідних клітин і неклітинних елементів, які мають спільне походження, будову та функції.
В організмі тварин та людини розрізняють чотири типи тканин, зокрема:
— епітеліальну;
— сполучну;
— м’язову;
— нервову.
Тканини тварин і людини вивчає гістологія. Серед тканий рослин виділяють малодиференційовані — твірні та високоди- ференційовані — постійні. Твірні тканини (меристема) забезпечують ріст, розвиток внутрішньої структури, формують постійні тканини, що становлять у рослинах системи: покривних тканин, провідних тканин, провітрювальних тканин і основних тканин. До системи основних тканин відносять механічні тканини рослин, асиміляційні (Хлоренхіма), запасаючі тканини, секреторні, або видільні (залозисті тканини рослин), та всисні тканини. Тканини рослин вивчає анатомія рослин.
Основні тканини (паренхіма)— це тканини рослин, що складаються із живих клітин різної форми і виконують різні функції— виповнюючу, асиміляційну, газообмінну, запасаючу, видільну та ін.
Гуморальні фактори взаємодії.
гуморальний фактор, стимулює антитілоутворення на піку імунної відповіді. Більшість вивчених медіаторів клітинного та гуморального імунітету виробляється лімфоцитами під впливом антигенних або мітогенних стимуляторів і є продуктами Т-клітин (Bloom, 1971; Pick, Turk, 1972; Waksman, Hamba, 1976).
З відомих чинників один лише тимозин виділяється епітеліальними клітинами тимуса в процесі нормального метаболізму і виконує найважливішу імунологічну функцію становлення Т-системи імунітету. Фактор, що забезпечує збільшення кількості зрілих антітелопродуцентов, синтезується клітинами кісткового мозку при відсутності будь-якої стимуляції. Методом диференціального центрифугування в градієнті щільності бичачого сироваткового альбуміну було показано, що клітини, відповідальні за синтез фактора, концентруються в легких фракціях і є, мабуть, бластних формами. У важких фракціях були виявлені клітини, які надають інгібуючу дію на синтез антитіл в зрілих антітелопродуценти, проте в вихідної популяції клітин кісткового мозку їх супресивна атівность не виявлялася (Михайлова, 1977).
Гуморальний фактор кісткового мозку, Стимулюючий антитілогенез в продуктивну фазу, термостабілен. Його активність зберігається при нагріванні до 56 ° С протягом 30 хв і знижується на 30% при 80 ° С. Супернатант, отримані після культивування клітин кісткового мозку, проявляють стимулюючу активність аж до розведення 1: 16.
Подальше вивчення властивостей стимулюючого фактора кісткового мозку дозволить з'ясувати і інші його фізико-хімічні характеристики, а виділення активної субстанції в чистому вигляді може мати і практичне значення.
Гуморальні фактори протиінфекційного захисту людини являють собою різні білки, розчинні в крові і рідинах організму.
Вони можуть самі мати антимікробні властивості чи здатні активувати інші гуморальні і клітинні механізми проти інфекційного імунітету. До неспецифічних гуморальних факторів імунітету відносяться:
— циркулюючі в крові інтерферони — вони підвищують стійкість кліток до дії вірусів, перешкоджають з розмноженню в клітках;
— З-реактивний білок крові — утворить комплекси зі збудниками інфекції, викликаючи тим самим активізацію системи комплементу, а також деякі клітки імунної системи (фагоцити й ін.);
— білки системи комплементу — звичайно неактивні, але здобувають імунологічну активність під впливом інших факторів імунітету;
— лізоцим — фермент, що розчиняє клітинні стінки інфекційних мікроорганізмів;
— трансферін — перешкоджає розмноженню мікроорганізмів;
До специфічних факторів гуморального імунітету відносяться білки, виділювані клітками імунної системи при специфічній її активації (інтерлейкіни, специфічні антитіла різних класів і ін.)
Цілком розділити клітинний імунітет і гуморальний неможливо: в ініціації утворення антител беруть участь клітки, а в деяких реакціях клітинного імунітету важливу сполучну функцію виконують антитіла.
Гуморальна імунна відповідь (утворення антитіл) являє собою кульмінацію ряду клітинних і молекулярних взаємодій, що відбуваються у визначеній послідовності:
- T-клітки розпізнають антиген, представлений ним антигенпрезентирующими клетками, і в результаті переходять в активований стан;
- Tх-клітки ( хелперние T-лимфоцити ) взаємодіють з B-клітками, що презентують їм антигенні фрагменти;
- активовані B-лімфоцити проліферують і диференціюються в антитілоутворюючі клітки;
- починається синтез антитіл і від їхнього класу залежить характер наступної імунної відповіді.
Поняття про тотипотентність клітин.
Тотипоте́нтість (омніпотентність) — здатність однієї клітини багатоклітинного організму давати початок цілому новому організму шляхом поділу. На відміну відплюрипотентних клітин, які можуть диференціюватись у будь-який із типів клітин дорослого організму, тотипотентні мають також здатність утворювати і позазародкові тканини, зокрема плаценту у ссавців. Тільки зигота та, у деяких організмів, її прямі нащадки (наприклад клітини морули) є тотипотентними[1]. У рослин соматичні клітини за певних умов, наприклад при обробці великими дозамиауксинів, можуть набувати здатності утворювати ембріон, тобто ставати тотипотентними[2].
Людські тотипотентні ембріональні стовбурові клітини можуть використовуватись для клітинної та генної терапії, а також тканинної інженерії з метою вирощування органів для трансплантації та заміни ушкоджених клітин. Таким чином вони можуть стати вирішенням проблеми багатьох захворювань, для яких зараз немає ефективного лікування. Проте використання ембріональних тотипотентних клітин пов'язане із певними етичними проблемами і має багато противників, через те що такі клітини можуть давати початок цілому організму
Тотипотентність (від лат. totus – весь, цілий, potencia – сила) – властивість клітин реалізувати власну генетичну інформацію, яка забезпечує їх диференціацію і розвиток до цілого організму. Тотипотентність клітин складає основу наукової ідеології біотехнології рослин. Такі методи біотехнології рослин як мікроклональне розмноження, соматичний ембріоїдогенез, культура зародків і гаплоїдів, соматична гібридизація і генна інженерія пов’язані з реалізацією тотипотентності клітин рослин трьома шляхами:
формування зачатків коренів або стебла бепосередньо з ексланта чи клітини;
формування зародкоподібних структур бепосередньо з ексланта чи клітини;
формування зачатків коренів,стебла або зародкоподібних структур через калусну тканину.
Калусна тканина – це маса неспеціалізованих клітин, які утворюються в результаті пошкодження або при вирощуванні експланта на штучному поживному середовищі.
Регенерація, гіпертрофія та атрофія тканин
Гіпертрофія (грец. hурег - над, понад, trophe - харчування) - збільшення об'єму тканини або органу внаслідок зміни розміру окремих клітин при збереженні їх кількості. Гіперплазія (грец. р1asis - освіта) - збільшення об'єму тканини або органу за рахунок збільшення кількості клітин.
Гіпертрофія буває істинна і помилкова. Справжня ги-пертрофія - збільшення обсягу тканини або органу і вище-ня їх функціональної здатності внаслідок розростися-ня паренхіматозних клітин, а також і інших елементів.
Прикладом є гіпертрофія гладеньких м'язів матки у вагітних тварин і збільшення залізистих клітин в лактаційний період, а також гіпертрофія серця при фізичній роботі. Хибна гіпертрофія - збільшення обсягу органу при розростання сполучної або жирової тканини.
Кількість паренхіматозних клітин при цьому залишається без змін або навіть зменшується, а функціональна здатність органу може знижуватися: наприклад, збільшення обсягу щитовидної залози при ендемічному зобі в результаті розростання її сполучної тканини або гіпертрофія молочної залози за рахунок жирової тканини.
Патологічна гіпертрофія - результат впливу несприятливих-приємних умов, що ведуть до надмірне навантаження органу: наприклад, виникнення робочої гіпертрофії серця у зв'язку із змінами його клапанного апарату або внаслідок гіпертонії, при емфіземи легенів та ін
Посилена робота і виникає рефлекторна порушення механізмів, що підвищують функціональну спроможність органу, його кровообіг, харчування та трофіку, обумовлюють гіпертрофію.
Розрізняють декілька видів гіпертрофії.
Робоча гіпертрофія. Спостерігають при підвищеній функціо-нальної навантаженні. Крім фізіологічних процесів
(тренінг коней тощо), можуть бути і патологічні, пов'язані з виникненням різних перешкод, які доводиться долати працює органу
(при звуженні стравоходу, кишечнику, сечовивідних шляхів, при вадах клапанів серця).
Замісна (компенсаторна), або вікарний, гіпертрофія можлива після видалення одного із парних органів. , Що залишився, орган, як правило, збільшується в обсязі: наприклад, гіпертрофія нирки, надниркових залоз, гіпертрофія однієї частки печінки при атрофії інший, гіпертрофія однієї частки легені при патології іншої.
У непарний органі, особливо лрі патології частини органу, може також виникнути компенсаторна гіпертрофія.
Прикладом може бути гіпертрофія кровотворної тканини (кісткового мозку, лімфатичних вузлів) при ураженні органа кровотворення - селезінки.
Компенсаторна гіпертрофія часто поєднується з гіперплазією клітин, що спостерігають при патології як парних, так і непарних органів.
Регенераційні гіпертрофія характеризується тим, що клітини ураженого органу починають посилено розмножувати ся (гіперплазія). Відзначають це після видалення частини органа (печінки, підшлункової залози, селезінки).
Кореляційна гіпертрофія виникає в органах, функціонально пов'язаних між собою; наприклад, після видалення щитовидної залози відбувається збільшення гіпофіза.
Робоча, замісна і регенераційні гіпертро-фії мають компенсаторне значення. Під впливом різних факторів процеси гіпертрофії можуть включатися відносно швидко. Виражений тканинний зростання вже спостерігають за кілька діб. Відзначається збільшення числа мітозов.В залежно від особливостей органу та існуючих умов термін появи максі-бітної гіпертрофії у тварин настає на 20 - 180-й день.
Експерименти показали, що суттєво на розвиток гіпертрофії впливають характер годування тваринного і діяльність регулюючих систем. Наприклад, значна компенсаторна гіпертрофія нирки наголошується в результаті підвищеного вживання білка в кормі.
У деяких випадках механізмом процесу гіпертрофії, мабуть, можуть бути нервово-ендокринні впливу.
Мається на увазі гіпертрофія здорової нирки при пошкодженні інший (пієлонефрит). Однак при недоліку соматотропного гормону передньої частки гіпофізу або при тіреоідектоміі компенсаторна гіпертрофія нирки Задерей жива.
Регенерація - відновлення тканин, органів, окремих частин живих істот, що зазнали руйнування або втрати. У процесі еволюційного розвитку в організмі виникла відповідна пристосувальна реакція.
Вона має велике значення не тільки для здорового організму, але і для хворого.
Розрізняють регенерацію фізіологічну та патологічну.
Фізіологічна регенерація - постійне відновлення відмерлих клітин молодими. Цей процес відбувається протягом усього життя організму. Інтенсивні процеси регенерації виявляють при відновленні клітин крові, епідермісу слизових оболонок. До них відносять також линьку ссавців і птахів, зростання різців у гризунів та ін Патологічна регенерація - процес відновлення пошкоджених тканин і органів після дії надзвичайних подразників.
Тканинна регенерація. У тварин, що стоять на нижчому щаблі розвитку, процеси йдуть більш повно.
У ссавців регенерувати можуть клітини всіх чотирьох видів тканин, але в різному ступені. Великий регенераторної здатністю володіють менш диференційовані тканини, ніж високодиференційовані. У регенерації тканин також має значення стадія онтогенетичного розвитку організму.
Сільськогосподарська здатність виявляється більш значною в ембріональному період життя.
Регенерація сполучної тканини. Пухка сполучна тканина має особливо високою здатністю до регене-рації. Найбільш активно регенерують інтерстиціальні елементи, що дуже важливо при запаленні і при ранового процесу. У зтіх випадках йде освіта сполучно-тканинної рубця, який заміщає виникає дефект.
Кісткова тканина також здатна до вираженої регенерації.
Основними елементами, відновлюють пошкоджений-у кісткову тканину, є остеобласти (малодіффе-ційованої камбіальні клітини кісткової тканини) .
Активно регенерують сухожилля, фасції, менш активно
- хрящі (регенерують не клітки хрящової тканини, а перихондрит) і дуже слабо - жирова тканина.
Регенерація епітеліальної тканини. Вираженої здатністю до регенерації володіють епітеліальні тканини - багатошаровий плоский епітелій шкіри, рогова оболонка ока. В процесі загоєння ран важливе значення має регенерація епідермісу. Відзначається висока Регенераторна здатність епітелію слизової оболонки в порожнині рота, губ, носа, шлунково-кишкового тракту, сечовому міхурі та ін Процеси регенерації віз-можна в паренхімі печінки, нирок, слинних залоз та ін
При наявності дратівливих факторів, значно підсилюють процеси регенерації, зростання епітеліальної тканини стано-вітся патологічним. Можуть виникнути атипові розростання епітелію і утворення в різних органах злоякісних пухлин.
Регенерація м'язової тканини виражена значно слабкіше, ніж тканин епітеліальної і сполучної.
Поперечно м'язові волокна регенерують амітотіческого шляхом ділення клітин здорової тканини, грані-чащіх з пошкодженим ділянкою.
Надалі що утворилися на кінцях пошкодженої м'язи своєрідні колбовідние випинання (м'язові нирки) зливаються.
У результаті відбувається відновлення поперечної ісчеренності у пошкоджених м'язових волокнах.
Гладка мускулатура відносно слабо регенерує і може відбуватися завдяки мітотичного поділу гладкоми-шечних клітин.
Регенерація нервової тканини. Нервова тканина володіє поганою здатністю до регенерації. Експерименти показали, що нервові клітини периферичної та вегето-нормативної нервової системи, рухові і чутливі нейрони спинного мозку досить слабко піддаються регене-рації. Навпаки, аксони нервових клітин (виключаючи кору, підкіркові вузли) регенерують краще. Крім того, в процесі відновлення нерва істотне значення мають шваннівською клітини (леммоціти). Вони формують своєрідні трубочки, в які і вростають регенері-ючий волокна центрального кінця повреіденного (перерва-занного) нерва. У мозку, де роль шваннівською клітин замінена клітинами глії, регенерація нервових волокон відсутня.
Атрофія - зменшення об'єму клітин відповідної тканини, органу і зниження їх фізіологічеської функції внаслідок недостатнього харчування або порушення обміну речовин. Гіпоплазія - недоразвігіе тканини, органа, частини тіла або цілого орга-нізма (в основі - порушення ембріогенезу).
Фізіологічна атрофія - зменшення обсягу окремих органів, тканин і ослаблення їх функції протягом нормального життя організму. Прикладом може бути інволюція зобної залози у дорослих тварин, матки у тварин після вагітності, пульпи молочних зубів з наступним їх випадінням.
Стареча атрофія. У результаті уповільнення процесів обміну речовин поступово слабшають і знижуються всі функції організму. Перш за все це відзначають високодиференційованих клітинах (зниження функції вищої нервової діяльності та паренхіматозних органів).
Значні зміни відбуваються в статевих залозах; наприклад, в яєчниках сповільнюється, а потім і зовсім припиняється дозрівання фолікулів, в насінниках - сперміїв; тварини стають безплідними. Знижуються кровотворна і регенеративна функція, слабшають кістково-м'язова система, функція серцево-судинної системи та інших органів, зменшується продуктивність тварин.
Атрофія від бездіяльності розвивається при зниженні діяльності органу, тканини протягом тривалого часу або при повному її відсутності. У результаті орган або тканину зменшується в розмірах. Спостерігається при денервації органу (параліч, парез), в експерименті при перерезке сухожилля, при тривалих і сильних больових роздратуваннях, а також якщо тварин містять в умовах значного обмеження їх рухової активності.
Атрофія від тиску. У результаті тривалого здавлення органу або тканини порушуються кровообіг і обмін речовин: наприклад, витончення альвеолярних перегородок при емфіземи легенів; атрофія паренхіми нирок при закупорки сечоводу і тиску сечі на тканину з боку балії (гідронефроз); атрофія тканини при стисненні її опухолио, стороннім тілом. неврогенні атрофія виникає в денервірованом органі чи тканині. Порушуються їх кровопостачання та іннервація.
До зтому виду відносяться, наприклад, недостатній розвиток м'язів, що покривають лопатки у коней ( «суха» лопатка), внаслідок порушення трофічної функції нервової системи, атрофія кінцівок, сім'яників, ослаблення росту у птахів при пошкодженні відповідних нервів.
Атрофія від ендокринної недостатності проявляється при порушенні функції ендокринної системи. При дисфункції аденогіпофіза може розвиватися виснаження організму - кахексія, а при гіпофункції щитовидної залози - тире-привносячи кахексія. В результаті порушення ендокринної функ-ції статевих залоз можливі місцеві атрофічні процеси, наприклад атрофія передміхурової залози після кастрації, атрофія матки і молочних залоз при видаленні яєчників.
Загальна характеристика епітеліальних тканин.
Епітеліальні тканини отримали свою назву від двох грецьких слів: «епі» - над і «тілі» - «сосочок». Вперше цей термін був застосований Рюішем, який назвав епітелієм тканину, розташовану над сосочками сполучної тканини, тобто покривний епітелій шкіри. Пізніше до групи епітеліальних стали відносити і інші прикордонні тканини, а потім і залози. Епітеліальні тканини утворені клітинами, званими епітеліоцитами.
Загальні морфологічні ознаки епітеліїв, що відрізняють їх від інших тканин, наступні:
1. Епітелій - це прикордонна тканина, яка відмежовує організм від зовнішньої або внутрішніх середовищ і одночасно здійснює з нею зв'язок. На поверхні організму перебуває покривний епітелій. Просвіти кишечника, незамкнуті порожнини нирок, матки та інших внутрішніх органів та їх проток також вистилають епітелієм. Нарешті, замкнуті порожнини - перикардіальна, плевральна і черевна - теж обмежені епітеліальної тканиною. Паренхіма внутрішніх органів у більшості випадків також складається з епітеліїв. Прикордонна функція є найважливішою ознакою епітеліальних тканин, що визначає структуру всіх їх різновидів.
2. Клітини епітеліїв розташовані у вигляді суцільних пластів, що покривають великі поверхні. Здатність утворювати пласти зберігається у епітеліїв і при вирощуванні їх в культурі тканин поза організмом. Розподіляючись пластом і функціонуючи як єдиний пласт, епітелій надійно відмежовує підлеглі тканини від зовнішнього для них середовища.
3. Основну масу епітеліальної тканини складають клітини, на відміну, наприклад, від сполучної тканини, де над клітинами значно переважають міжклітинні структури. Багатство клітинами також визначається прикордонним становищем епітеліїв.
Слід зазначити, однак, що деякі епітелії виробляють досить потужний шар міжклітинної речовини у вигляді кутикулярних структур, які за площею можуть переважати над епітеліоцитами. При цьому епітелій зберігає характер пласта, має тісне розташування клітин, а кутикула відкладається на вільній поверхні, як це спостерігається у покривах багатьох безхребетних тварин. Між тісно розташованими епітеліальними клітинами знаходиться дуже мало міжклітинної речовини. На думку ряду авторів, воно між сусідніми клітинами взагалі відсутня, а щілини між клітинами шириною близько 20 нм заповнені глікокалік-сом. Спочатку глікокалікс був виявлений на поверхні мікроворсинок ентероцитів, а в даний час - і на поверхні інших епітеліальних і неепітеліальних клітин.
Прикордонне положення епітеліїв і в зв'язку з цим часта їх вразливість привели в процесіеволюції до вироблення у них високої здатності до регенерації у відповідь на пошкодження.Регенерація епітелію є результатом розмноження або гіпертрофії клітин.
4. Епітелії, особливо одношарові, характеризуються різко вираженою полярністю утворюють їх клітин, тобто базальна і апикальная частини епітеліоцитів значно відрізняються один від одного і структурно, і функціонально. Так, епітеліальні клітини кишечника мають на своїй апікальної поверхні мікроворсинки; в над'ядерний області розташовано комплекс Гольджі, тоді як у базальних частинах клітин немає ні мікроворсинок, ні КГ. Через апикальную зону клітини всмоктуються продукти травлення, а через базальну частину здійснюється її живлення і виділення в кров і лімфу продуктів всмоктування. У багатошарових епітелію також спостерігається полярність шару клітин.
5. Епітелій завжди розташовується на чітко вираженої базальної мембрані, тобто на шарі міжклітинної речовини, утвореного діяльністю клітин як епітелію, так, можливо, і підлягає сполучної тканини. Базальна мембрана знаходиться тільки з базальною боку клітин і зміцнює епітеліальний пласт, сприяючи виконанню ним своєї основної прикордонної функції. Крізь базальну пластину здійснюється харчування епітелію. Слід підкреслити, що кровоносних і лімфатичних судин в епітелії немає. Проникнення деяких речовин із зовнішнього середовища в підлягає сполучну тканину також відбувається через епітелій. Таким чином, базальна пластина, відокремлюючи епітелій від сполучної тканини, одночасно зв'язує їх між собою в епітеліальної-сполучнотканинний комплекс.
Незважаючи на схожість епітеліїв по перерахованих ознаками, кожен вид епітелію має свої індивідуальні особливості.
Види епітеліїв.
Епіт́елій, епітеліальна тканина (лат. epithelium, від грецького епі- +thele — молочна залоза, синонім — епітеліальна тканина) — шар клітин, що вистилає поверхню (епідерміс) і порожнини тіла, а також слизисті оболонки внутрішніх органів, харчового тракту, дихальної системи,сечостатеві шляхи. Крім того, утворює більшість залоз організму. Складається з клітин, які щільно розташовані одна біля одної, міжклітинної речовини мало.
Види епітелію:
покривний
залозистий
Морфологічна класифікація покривного епітелію залежить від кількості шарів та форми клітин. Тому, перш за все, епітелій поділяють на одно- та багатошаровий.
В одношаровому епітелії всі клітини розташовані на базальній мембрані. У багатошаровому на базальній мембрані лежить лише нижній базальний шар епітеліоцитів, а клітини, що розташовані вище, втрачають контакт із нею. Одношаровий епітелій поділяють на однорядний та багаторядний, а однорядний, в свою чергу, за формою клітин буває плоским, кубічним, призматичним. В однорядному епітелії всі клітини мають однакову форму і розмір, а їх ядра розташовуються на одному рівні. У багаторядному епітелії внаслідок різних видів, розмірів і форм клітин їх ядра лежать на різних рівнях, але всі епітеліоцити контактують з базальною мембраною.
Залозистий епітелій складається із залозистих клітин – гландулоцитів. Це спеціалізовані клітини, що пристосовані до синтезу, накопичення і виведення секрету.
Розрізняють залози внутрішньої секреції – ендокринні і залози зовнішньої секреції – екзокринні, відповідно, гландулоцити поділяють на ендокриноцити та екзокриноцити. Ендокринні залози становлять ендокринну систему, яка разом із нервовою системою регулює і координує роботу органів усього організму. Характерним для них є те, що свої секрети (гормони) вони виділяють безпосередньо в кров. До таких залоз належать гіпофіз, епіфіз, щитоподібна і прищитоподібні залози, надниркові залози, ендокринні відділи підшлункової та статевих залоз.
Функції:
захисна (епітелії шкіри)
секреція (епітелій, що входить до складу залоз)
всмоктування (епітелій шлунково-кишкового тракту)
виділення (епітелій органів виділення)
газообмін (епітелій легеневих пухирців)
Головні особливості епітеліальних тканин — швидка регенерація і відсутність кровоносних судин.
Поділяється на одношаровий, багатошаровий, залозошаровий, війчастий і залозистий епітелій, а також розрізняють види за формою клітин тканини: кубічний, плоский, циліндричний.Одношаровий епітелій вкриває стінки судин і внутрішніх органів, бере участь в обміні речовин та частково виконує захисну функцію. Багатошаровий — це зовнішній покрив людини, тобто шкіра. Відмежовує зовнішнє середовище від внутрішнього, захищає від механічних пошкоджень та проникнення мікроорганізмів. Багатошаровий епітелій найшвидше відновлюється, вистилає порожнину рота. Війчаста епітеліальна тканина вистилає носову порожнину, затримує пил та різні частинки, вистилає й тонкий кишечник, де виконує всмоктувальну функцію. Залозистий входить до складу залоз і забезпечує виділення гормонів і секретів.
Морфо-функцїональна та філогенетична класифікація епітеліальних тканин.
У межах епітеліальних тканин існує досить велика різноманітність клітинних типів.Залежно від мети, яку ставить перед собою дослідник, що вивчає цю тканину, можна класифікувати епітелії за структурою, функціональним особливостям, походженням, положенню епітелію по відношенню до середовища, з якою він межує, за здатністю до оновлення та іншими ознаками. Відповідно використовуються морфологічна, онто-і філогенетичні, функціональна класифікації, а також класифікації за проліферативним потенціям і властивостями прикордонне ™.
Морфологічна класифікація. За морфологією можна розділити епітелії на кілька різновидіввідповідно до взаємним розташуванням клітин у складі пласта і їх формою. Якщо клітини в пласті розташовані в один шар - епітелій називається одношаровим, а якщо в кілька шарів - багатошаровим. Зустрічаються також багаторядні епітелії, у яких усі клітини лежать на базаль-ної мембрані, як в одношаровому епітелії, але лише частина їх апікальних решт доходить до вільної поверхні. Вершини інших, більш низьких клітин, виявляються між бічними поверхнямисусідніх клітин. Таким чином, в багаторядним епітелієм клітини мають різну висоту і форму: одні - циліндричну, а інші - пірамідальну. Такі клітини називають Інтернейрони.
У ряді випадків, однак, було показано, що в деяких, які вважалися багаторядними, епітелію в дійсності є однослоіность, тобто вершини всіх клітин доходять до вільної поверхні, а Інтернейрони клітини мають форму не однієї, а двох пірамід, з'єднаних вершинами. Таку форму клітин можна побачити тільки на ультратонких, суворо орієнтованих зрізах в електронний мікроскоп.
Серед багатошарових епітеліїв виділяють зроговілий і неорогове-вающие. Своєрідним різновидом багатошарового епітелію є перехідний епітелій. Кількість шарів в ньому міняється залежно від ступеня наповнення органу, порожнину якого він вистилає. Ця класифікація, хоча широко застосовується, не дає уявлення про функції та походження різних епітеліїв.
Функціональна класифікація. Найбільш повну характеристику епітеліальних тканин можна отримати, користуючись функціональної класифікації. Однак вона має той недолік, що функції епітеліїв вельми
У більшості тварин можна виділити наступні типи епітеліїв за належністю їх до тих або іншим органам, які мають своєрідними функціональними ознаками:
1. Покривний епітелій.
2. Епітелій шлунково-кишкового тракту.
3. Епітелій дихальних шляхів.
4. Нирковий епітелій.
5. Мезотелий.
6. Епітелій гонад.
7. Залозистий епітелій і ін
У кожній з цих груп епітеліїв, у свою чергу, можна виділити радий різновидів.Наприклад, в епітелії нирок є і подоцитів, що утворюють внутрішній листок боуменовой капсули, і плоский, кубічний і циліндричний епітелії ниркових канальців, кожний з яких має своєрідну будову, функції і т.д. Ці різні типи клітин притаманні, однак, одному органу і в сукупності один з одним і іншими тканинами органу здійснюють загальний секреторний та осморегулюючі; ефект. Недоліком цієї класифікації є те, що призначення і структура різних типів епітеліоцитів в одному органі часто бувають більш різноманітними, ніж епітеліальних клітин різних органів. Наприклад, епітеліоцити проток різних органів мають між собою дуже багато спільного і різко відрізняються за структурою та функціями від паренхіматозних клітинвідповідних органів.
Робляться спроба розділяти епітелії по функціях конкретних клітин, але за такої класифікації пропадає характеристика епітелію як тканини.
Онто-і філогенетичні класифікація. Класифікація епітеліїв за ознакою їх походження з певних зародкових листків і закладок, а також за своєрідним характером росту в культурі тканин була запропонована Н.Г. Хлопіна. Він розділив епітелії хребетних на наступні типи:
- Епідермальний, що виникає з ектодерми;
- Ентеродермальний, що стався з ентодерми;
- Целонефродермальний, що утворився з мезодерми;
- Епендімогліальний, розвивається із зачатка нервової трубки;
- Ангіодермальний, що має мезенхімного походження.
Частина дослідників, однак, останні два різновиди тканини не відносять до епітелію.Вистилку спинномозкового каналу і шлуночків мозку - епендима - вони відносять до нервової тканини, а вистилку кровоносних судин - ендотелій - до тканин внутрішнього середовища.
В.П. Михайлов, виходячи з уявлення про те, що джерелами розвитку тканин певного типу бувають не зародкові листки, а однаковим чином детерміновані комплекси клітин незалежно від їх формального розташування в тому чи іншому зародковому листку, запропонував наступну класифікацію тканин. В один тканинної тип В.П. Михайлов об'єднав різні тканини, але утворені із загальних зачатків і здатні в ряді умов переходити один в одного.
Класифікація тканин, запропонована В.П. Михайловим, має ряд недоліків. Зокрема, ембріональними зачатками в ній називаються епідерміс, епітелій целома, епітеліальна вистилання середньої кишки та її похідні. Ці структури, як вказує А.А. Кліше, навряд чи доцільно відносити до ембріональних зачатків. Крім того, у графу «конкретні тканини» занесені не тільки тканини, а й клітини і органи.
А.А. Кліше, розробляючи проблему класифікації тка-1 ній, вважав, що вона повинна відображати стійку детермінованість четирехтканевих систем, кожна з яких включає велику кількість груп! і різновидів тканин, що виникли на основі дивергентной еволюції. 1 Вихідним і головним критерієм при виявленні різновидів тканин автором був узятий принцип їх детермінованості, наслідком якої, є нездатність до перетворень тканин один в одного і стійке збереження ними своїх гістобластіческіх потенцій. У системі епітеліальних] тканин А.А. Кліше виділяв п'ять типів:
1. Епітелії шкірного типу.
2. Епітелії кишкового типу.
3. Епітелії ниркового типу.
4. Епітелії целомического типу.
5. Епітелії гліального типу.
При конкретному розподіл тканин по групах, однак, виникає ряд неузгодженостей, так до групи епітеліїв шкірного типу автор відносить настільки несхожі по структурам і функцій тканини, як епідерміс, епітелій щитовидної залози і альвеолярний епітелій. Тут за основна ознака було вибрано походження тканин. З іншого боку, епітелій нирковий і целомічних розглядаються як окремі типи, незважаючи на їх генетичну близькість. Суперечливим є відносити до епітелію епендімнуюглію, а до гліальними типу - смаковий епітелій.
Ще одну класифікацію епітеліїв, названу авторами комплексної, пропонують І.М.Борисов і співавт. У ній враховується як походження тканинних різновидів, так і їх специфічна органна приналежність. У цій досить вдалою класифікації не надто виправдано виділення авторами в окрему групу вагінального епітелію, а епітелій нирок і целомічних недоцільно розглядати, як окремі типи, незважаючи на їх генетичну близькість.
Існує ще цілий ряд класифікації епітеліїв з урахуванням їх проліферативної активності, здатності до регенерації і ін Кожна з наведених класифікацій має право на існування, особливо, якщо тканини вивчаються і описуються під певним кутом зору. Наприклад, досліджуютьсяпроцеси їх розвитку, регенераторні здібності, їх проліферативний пул і т.д., але тільки морфофункциональная класифікація дає найбільш загальну і істотну характеристику клітин в тканинних системах.
Всі інші класифікації тканин мають більш приватний характер.
Особливості будови епітеліоцитів.
Одиночні епітеліоцити в ході распластиванія набувають дискоїдний форму і не рухаються направлено по субстрату. Псевдоподіальная активність в таких клітинах розподілена по всьому периметру.
Псевдоподіальная активність в епітеліальних острівці або в моношарі, загоювати рану, розподілена по зовнішньому периметру, на ділянках краю, не утворюють контакту з сусідніми клітинами. Таким чином, псевдоподіальная активність епітеліальних клітин не є поляризованої в якомусь певному напрямку.
Епітеліоцити: міжклітинні контакти
Характерною особливістю епітеліальних клітин є їх здатність утворювати великі і міцні міжклітинні контакти.
Безліч епітеліоцитів, що контактують один з одним, формують монослой зчеплених між собою клітин. У складі клітинних пластів епітеліальні клітини виявляються поляризованими щодо базоапікальной осі (Rodriguez-Boulan E., Nelson WJ, 1992; Rodriguez-Boulan E., Powell SK, 1992).
Клітинна поверхня розділяється на апікальну і базо-латеральну за допомогою адгезійних поясів, смуги міжклітинних контактів, оперізують кожну клітину в складі епітеліального пласта або острівця.
Епітеліоцити: організація актину
При флуоресцентної забарвленні на актин в епітеліальних клітинах виділяється кілька видів структур.
Головна особливість організації актину в епітеліальних клітинах – кільцевої пучок актину. В одиночних епітеліальних клітинах з псевдоподіальной активністю, розподіленої по всьому клітинному периметру, такий пучок замкнутий у кільце. У периферичних клітинах епітеліального острівця або на краю монослоя, що мають псевдоподіальную активність лише в тих ділянках клітинного краю, які не утворюють контактів з сусідніми клітинами, такий пучок має вигляд незамкненою дуги.
Ще один тип структур – прямі пучки Актинові філаментів, стрессфібрілли, які містяться в цитоплазмі під різними кутами до краю клітки і закінчуються фокальними контактами з субстратом, в яких виявляються вінкулін та інші специфічні білки. Стрессфібрілли маються на одиночних дискоїдний епітеліоцитах і в клітинах, організованих в моношар.
Міжклітинні контакти адгезійного типу асоційовані з Актинові філаментах, тому при флуоресцентному фарбуванні на актин ці контакти виглядають як тонкий обідок навколо клітини в апікальній частині (Geiger B. et al., 1987).
В одиночних епітеліоцитах, в крайових клітинах епітеліального острівця або пласта, що рухається в рану, де є вільний край з псевдоподіальной активністю, актинових цитоскелет має зональну організацію (Світкіна Т.М., 1989). Самою дистальної є зона активного краю, що містить густу мережу мікрофіламентів і примикає до неї зону розрідження.
Наступною в напрямку від периферії до центру клітини є зона власне ламелли. Вона містить прямі пучки Актинові філаментів або стрессфібрілли. Центром організації мікротрубочок культивованих епітеліоцитів є центросома, локалізована близько ядра. Від неї цитоплазматичні мікротрубочки розходяться радіально до клітинної периферії, де або закінчуються, або згинаються і повертають. В результаті функціональної базоапікальной поляризації епітеліальних клітин в ході утворення епітеліальних пластів розподіл мікротрубочок змінюється (Bacallao R. et al., 1989; Rodriges-Boulan E., Nelson WJ, 1989).
В ході поляризації центросоми зміщуються до апікальній мембрані, мікротрубочки виявляються організованими в мережу, розташовану в апікальній частині клітини, і пучки мікротрубочок, витягнутих уздовж базолатеральной осі клітини, мінус і плюс кінці яких розташовані в апікальній і базальній частинах клітин відповідно.
Епітеліоцити: трансформація: зміна фенотипу клітин
Заміщення комплексу ознак, властивого в нормі нерухомим епітеліальних клітин, фенотипом, властивим направлено двигающимся клітинам може відбуватися під впливом низки чинників. Наприклад, показано, що під дією цитокіну HGF / SF нормальні епітеліальні клітини втрачають контакти з сусідами, поляризуються і набувають здатність направлено рухатися (Dowrick PG et al., 1991; Prescott AR et al., 1992; Stoker M. et al., 1987 ).
При распластиванія на малоадгезівном або неоднорідно адгезивному субстраті, в нормі дисковидні, добре розпластані клітини лінії IAR-2 поляризуються, а ступінь їх распластанность істотно знижується (Domnina LV et al., 1995).
Епітеліальні клітини, піддані трансфекції набувають фібробластоподібних поляризований фенотип (Behrens J. et al., 1989).
Епітеліоцити лінії IAR-2 стійко екпрессірующіе екзогенний онкоген ras, втрачають дисковидні форму, набуваючи фібробластоподібних поляризований фенотип, ступінь їх распластанность зменшується (Gloushankova NA et al., 19; Gloushankova NA et al., 1995).
При пухлинної трансформації морфологія культивованого епітелію змінюється: відбувається погіршення распластиванія клітин, порушення міжклітинних контактів, витягування і поляризація клітин (Гельштейн В.І., 1981; Vasiliev JM, Gelfand IM, 1981).
Подібні зміни зручно вивчати, використовуючи клітинні лінії, одного походження, які мають різну пухлинну прогресію, будучи трансформованими спонтанно або за допомогою канцерогенних речовин. Прикладом такої серії є тривало культивовані лінії клітин щурячої печінки серії IAR, отримані Монтесано (Montesano R. et al., 1977; Montesano R. et al., 1975; Bannikov GA, Saint-Vincent L., Montesano R., 1980).
У залежності від ступеня трансформації, клітини послідовно втрачають ознаки, властиві епітелію в нормі (Kaighn ME et al., 1980). На початкових етапах пухлинної прогресії слабшають міжклітинні контакти, клітини набувають здатність рости поодинці, при цьому зберігаючи дискоїдний форму і псевдоподіальную активність, розподілену по всьому периметру клітини. На наступних етапах з’являються стабільні ділянки краю, частка яких по відношенню до псевдоподіально активним ділянкам з зростанням ступеня трансформації клітин збільшується. Клітини при цьому набувають фібробластоподібних поляризований фенотип. Всі ці зміни відбуваються послідовно і супроводжуються зростанням опухолеродного потенціалу клітин (Montesano R. et al., 1975; Montesano R. et al., 1977).
Епітелій: поляризація
В епітеліальних клітинах MDCK крім звичайних мікротрубочок, асоційованих з центросомой і мають радіальне розташування, маються нецентросомальние мікротрубочки (Bre MH et al., 1987). Вони утворюють пучки, розташовані по периферії клітини і орієнтовані в базо-апікальному напрямку поляризованого епітелію. При цьому “+”-кінці направлені до базальної поверхні клітин, а “-”-кінці – до апікальної. Ці мікротрубочки характеризуються високою стабільністю, стійкістю до руйнування нокодазол ом і високим вмістом детірозінілірованного тубуліну. Аналогічні стабільні мікротрубочки виявляються і в інших типах епітеліальних клітин. Їх функцією є підтримка поляризованої форми і здійснення спрямованого руху органел (Gelfand VI, Bershadsky AD, 1991).
Міжклітинні з'єднання епітеліоцитів.
Характерной особенностью эпителиальных клеток является их способность образовывать обширные и прочные межклеточные контакты .
Множество эпителиоцитов, контактирующих друг с другом, формируют монослой сцепленных между собой клеток. В составе клеточных пластов эпителиальные клетки оказываются поляризованными относительно базоапикальной оси
Клеточная поверхность разделяется на апикальную и базо- латеральную при помощиадгезионных поясов , полосы межклеточных контактов, опоясывающих каждую клетку в составе эпителиального пласта или островка.
Подразделяются на два основных вида;
1. Механические соединения - Обусловливают механическую связь эпителиоцитов друг с другом. В их число входят Плотные соединения, промежуточные соединения, десмосомы, интердигитации;
2. Коммуникационные соединения - (от лат. communicatio - сообщение) обеспечивают химическую (метаболическую, ионную и электрическую) связь между эпителиоцитами. К ним относятся Щелевые соединения.
(1) Плотное соединение (Zonuia Occludens - поясок замыкания) - Наиболее тесный контакт клеток из всех известных в природе. Представляет собой область Частичного слияния наружных листков плазмолемм Двух соседних клеток, которая блокирует распространение веществ по межклеточному пространству (обеспечивая тем самым барьерную функцию эпителия и регулируемость транспорта веществ через эпителиальный пласт). Это соединение также препятствует свободному перемещению и смешиванию функционально различных внутримембранных белков, локализующихся в плазмолемме апикальной и базолатеральной поверхностей клетки, что способствует поддержанию ее полярности.
Плотное соединение имеет вид пояска шириной 0.1-0.5 мкм, окружающего клетку по периметру (обычно у ее апикального полюса) и состоящего из Анастомозирующих тяжей внутримембранных частиц. Эти частицы образованы белком Окклюдином; Каждая из них представляет собой область точечного слияния плазмолемм двух соседних клеток. Проницаемость плотных соединений тем ниже, чем выше число тяжей таких частиц. Для поддержания целостности этих соединений Необходимы двухвалентные катионы (Са2+, Mg2+). Они могут динамично перестраиваться (вследствие изменений экспрессии и степени полимеризации окклюдина) и временно размыкаться (например, при миграции лейкоцитов через межклеточные пространства).
(2) Промежуточное соединение, или опоясывающая десмосома '(Zonula Adherens - поясок Сцепления)Локализуется на латеральной поверхности эпителиоцита между областью расположения плотного соединения и десмосом (что обусловило его первое название). Охватывает клетку по периметру в виде пояска , на сечении имеющего сходство с десмосомой (что послужило основанием для второго названия). В области промежуточного соединения обращенные к цитоплазме листки плазмолеммы утолщены, образуя Пластинки прикрепления, Которые содержат Актин-свяшвающие белки а-актинин, винкулин и плакоглобин (последний обнаруживается также в десмосомах). К этим пластинкам прикрепляются элементы цитоскелета - Актиновые микрофиламенты, Вплетающиеся также в терминальную сеть. Межклеточная щель расширена до 15-20 нм и заполнена умеренно плотным веществом, в состав которого входит адгезивный трансмембранный гликопро-теин Е-кадгерин, Обеспечивающий в присутствии ионов Са2+ связь между соседними клетками. Со стороны цитоплазмы в области промежуточного соединения к Е-кадгерину через А-актинин и Винкулин ПрикрепляютсяАктиновые микрофиламенты, Что обусловливает Связь цитоскелета с компонентами межклеточного вещества.
(3) Десмосома (Macula Adherens - пятно сцепления) - Состоит из утолщенных и уплотненных участков питоплазматического листка плазмолемм двух соседних клеток - Пластинок прикрепления, РазделенныхМежклеточной щелью
Базальна мембрана та її функції.
Базальна мембрана, основна мембрана — тонкий шар волокон, що залягає під епітелієм (вистеляє порожнини і поверхні тіла) чи ендотелієм(вистеляє внутрішні поверхні кровоносних судин).
Базальну мембрану часто плутають з базальною пластинкою, яка є частиною базальної мембрани і була відкрита пізніше при розгляданні базальної мембрани під електронним мікроскопом. Фактично під поняттям базальної мембрани слід розуміти саме базальну пластинку. Під світловим мікроскопом можна побачити лише базальну мембрану.
Базальна мембрана (лат. membrana basalis) утворюється при зливанні двохшарової базальної пластинки (lamina basalis) і ретикулярної пластинки (lamina reticularis). Під світловим мікроскопом має вигляд однорідного шару. Базальна пластинка прикріплюється до ретикулярної пластинки за допомогою фіксуючих волокон колаген IV і мікрофібрил (фібрилін).[1]
Базальну пластинку в свою чергу можна поділити на два шари:
світла пластинка (lamina lucida) межує з епітеліальними клітинами;
темна пластинка межує (lamina densa) зі сполучною тканиною.
Під електронним мікроскопом темна пластинка має товщину 30-70 нм і складається з сітки колагенових волокон типу IV. Колагенові волокна оповиті перлеканом, який являє собою глікозаміноглікан гепарансульфат.[2] Світла пластинка утворена зламініну, інтегринів, енактину, дистрогліканів)
Для кращого розуміння будови базальної мембрани шари впорядковано слідуючим чином:
Епітеліальна тканина (ззовні)
Базальна мембрана
Базальна пластинка
Світла пластинка
ламінін
інтегрин
енактин
дистроглікани
Темна пластинка
колаген IV (сполучені з перлеканом, багаті на гепарансульфат)
Скріплюючі білки (між базальною і ретикулярною пластинкою)
колаген VII тип (скріплюючі волокна)
фібрилін
Ретикулярна пластинка
колаген III
Сполучна тканина (всередині)
Функції:
Основною функцією базальної мембрани є фіксація епітелію до нижче розміщеної сполучної тканини. Це досягається через клітинно-матриксну адгезію за допомогою адгезивних (скріплюючих) білків, присутніх в базальній мембрані.
Базальна мембрана діє як механічний бар'єр, попереджуючи на ранніх стадіях розповсюдження пухлин в більш глибокі тканини (карцинома in situ).[3]
Базальна мембрана має велике значення в ангіогенезі (утворення нових кровоносних судин).
Білки базальної мембрани можуть пришвидшувати диференціацію епітеліальних клітин.[4]
Через базальну мембрану проходить постачання поживних речовини епітеліальним клітинам, оскільки ті не мають власних кровоносних судин.
Епітеліальні клітини є поляризовані відносно базальної мембрани. Вони мають базальний (повернений в сторону базальної мембрани) і апікальний (повернений назовні) полюси.
Одношарові епітелії.
Одношаровий плоский епітелій ( ендотелій і мезотеліом). Ендотелій вистилає зсередини кровоносні, лімфатичні судини, порожнини серця. Ендотеліальні клітини плоскі, бідні органелами і утворюють ендотеліальний пласт. Добре розвинена обмінна функція. Вони створюють умови для кровотоку. При порушенні епітелію утворюються тромби. Ендотелій розвивається з мезенхіми. Другий різновид - мезотеліом - розвивається з мезодерми. Вистилає всі серозні оболонки. Складається з плоских полігональної форми клітин, пов'язаних між собою нерівними краями. Клітини мають одне, рідше два сплощені ядра. На апикальной поверхні є короткі мікроворсинки. Вони мають всмоктувальної, видільної та розмежувальної функціями. Мезотеліом забезпечує вільне ковзання внутрішніх органів відносно один одного. Мезотеліом виділяє на свою поверхню слизовий секрет. Мезотеліом запобігає утворенню сполучнотканинних спайок. Досить добре регенерують за рахунок мітозу.
Одношаровий кубічний епітелій розвивається з ентодерми і мезодерми. На апікальної поверхні є мікроворсинки, що збільшують робочу поверхню, а в базальної частини цитолемма утворює глибокі складки, між якими в цитоплазмі розташовуються мітохондрії, тому базальна частина клітин виглядає покреслений. Вистилає дрібні вивідні протоки підшлункової залози, жовчні протоки і ниркові канальці.
Одношаровий циліндричний епітелій зустрічається в органах середнього відділу травного каналу, травних залозах, нирках, статевих залозах і статевих шляхах. При цьому будова і функція визначається його локалізацією. Розвивається з ентодерми і мезодерми. Слизову шлунка вистилає одношаровий залозистий епітелій. Він виробляє і виділяє слизовий секрет, який поширюється по поверхні епітелію і захищає слизову оболонку від пошкодження. Цитолемма базальної частини також має невеликі складки. Епітелій має високу регенерацією.
Ниркові канальці і слизова оболонка кишечника вистелена каемчатая епітелієм. У каемчатая епітелії кишечника переважають каемчатая клітини - ентероцита. На їх верхівці розташовуються численні мікроворсинки. У цій зоні відбувається пристінкові травлення та інтенсивне всмоктування продуктів харчування. Слизові келихоподібних клітини виробляють на поверхню епітелію слиз, а між клітинами розташовуються дрібні ендокринні клітини. Вони виділяють гормони, що забезпечують місцеву регуляцію.
Одношаровий багаторядний Війчастий епітелій. Він вистилає повітроносні шляхи і має ентодермальні походження. У ньому клітини різної висоти, і ядра розташовуються на різних рівнях. Клітини розташовуються пластом. Під базальної мембраною лежить пухка сполучна тканина з кровоносними судинами, а в епітеліальних шарі переважають високодиференційовані війчасті клітини. У них вузьке підстава, широка верхівка. На верхівці розташовуються миготливі війки. Вони повністю занурені у слиз. Тим війчасті клітини знаходяться келихоподібних - це одноклітинні слизові залози. Вони виробляють слизовий секрет на поверхню епітелію. Є ендокринні клітини. Між ними розташовуються короткі і довгі вставні клітини, це стовбурові клітини, малодиференційовані, за рахунок них йде проліферація клітин. Миготливі війки здійснюють коливальні рухи та переміщують слизову плівку по повітроносних шляхах до зовнішнього середовища.
Багатошарові епітелії.
Багатошаровий плоский незроговілий епітелій. Він розвивається з ектодерми, вистилає роговицю, передній відділ травного каналу і ділянка анального відділу травного каналу, піхву. Клітини розташовуються в декілька шарів. На базальної мембрані лежить шар базальних або циліндричних клітин. Частина з них - стовбурові клітини. Вони проліферують, відокремлюються від базальної мембрани, перетворюються на клітини полігональної форми з виростами, шипами і сукупність цих клітин формує шар шипуватий клітин, розташованих у кілька поверхів. Вони поступово стають більш щільними і утворюють поверхневий шар плоских, які з поверхні відриваються в зовнішнє середовище.
Багатошаровий плоский зроговілий епітелій - епідерміс, він вистилає шкірні покриви. У товстій шкірі (долоні поверхні), яка постійно відчуває навантаження, епідерміс містить 5 шарів:
1 - базальний шар - містить стовбурові клітини, диференційовані циліндричні і пігментні клітини (пігментоціти).
2 - шипуватий шар - клітини полігональної форми, в них містяться тонофібрілли.
3 - зернистий шар - клітини набувають ромбоподібну форму, тонофібрілли розпадаються і всередині цих клітин у вигляді зерен утворюються білок кератогіалін, з цього починається процес зроговіння.
4 - блискучий шар - вузький шар, в ньому клітини стають плоскими, вони поступово втрачають внутрішньоклітинну структуру, і кератогіалін перетворюється на елеідін.
5 - роговий шар - містить рогові лусочки, які повністю втратили будова клітин, містять білок кератин. При механічного навантаження і при погіршенні кровопостачання процес зроговіння посилюється.
У тонкій шкірі, яка не відчуває навантаження, відсутня зернистий і блискучий шари.
Багатошаровий кубічний і циліндричний епітелії зустрічаються вкрай рідко - в області кон'юнктиви очі й області стику прямої кишки між одношаровим і багатошаровим епітелієм.
Перехідний епітелій (уроепітелій) вистилає сечовивідні шляхи і аллантоис. Містить базальний шар клітин, частина клітин поступово відокремлюється від базальної мембрани і утворює проміжний шар грушовидних клітин. На поверхні розташовується шар покривних клітин - великі клітини, іноді дворядні, покриті слизом. Товщина цього епітелію змінюється в залежності від ступеня розтягування стінки сечовивідних органів. Епітелій здатний виділяти секрет, що захищає його клітини від дії сечі.
Залозистий епітелій - різновид епітеліальної тканини, яка складається з епітеліальних залозистих клітин, які в процесі еволюції набули провідне властивість виробляти й виділяти секрети. Такі клітини називаються секреторними (залозистими) - гландулоцітамі. Вони мають таку саму загальну характеристику як покривний епітелій. Розташований в залозах шкіри, кишечнику, слинних залозах, залозах внутрішньої секреції і ін Серед епітеліальних клітин знаходяться секреторні клітини, їх 2 види.
екзокринні - виділяють свій секрет в зовнішнє середовище або просвіт органа.
ендокринні - виділяють свій секрет безпосередньо в кровотік.
Залозистий епітелій.
Залозисті клітини (гландулоцити) синтезують секрет і виводять продукти синтезу. Розрізняють ендокринні залози, що синтезують гормони і виділяють їх у кров, і екзокринні залози, які виводять продукти секреції на поверхню епітелію.
Екзокринні залози мають вивідні протоки і кінцеві секреторні відділи.
У залежності від локалізації виділяють залози ендоепітеліальні і екзоепітеліальні.
У залежності від будови вивідної протоки залози поділяють на прості і складні. Якщо вивідна протока розгалужена залози називаються складними, якщо нерозгалужена — простими.
У залежності від кількості секреторних відділів залози поділяють на розгалужені (мають кілька секреторних відділів) і нерозгалужені (мають один секреторний відділ).
У залежності від типу секреції розрізняють:
-мерокринові залози (виділяють секрет без порушення цілісності клітини),
-апокринові (апікальна частина клітини відокремлюється разом із секретом),
-голокринові (клітина цілком руйнується).
У залежності від форми кінцевого секреторного відділу залози бувають трубчасті, альвеолярні і альвеолярно-трубчасті.
Загальним для епітеліальної тканини є те, що вона складається з клітин, які щільно прилягають одна до одної, та незначної кількості міжклітинної речовини. Залежно від функції розрізняють покривний та залозистий епітелій. Покривний епітелій розміщується на всій поверхні тіла та на стінках внутрішніх порожнистих органів. Залозистий епітелій створює робочу частину залоз.
Залозистий епітелій, основна функція якого – секреція (виділення) різних речовин.
За походженням всі типи епітеліальної тканини походять від епітеліальної тканини походять від ектодерми (епідерміс шкіри, ротової порожнини, частини стравоходу та деяких інших поверхонь, що контактують із зовнішнім середовищем та можуть зазнавати механічного впливу) та ентодерми (епітелій внутрішніх ділянок шлунково-кишкового тракту, легень, залозистий епітелій тощо). Внутрішній шар серця та кровоносних судин, який дуже схожий на одношаровий плоский епітелій, але має мезодермальне походження, отримав власну назву – ендотелій). Вживають також поняття "складні епітелії" - багатошарові покривні утворення, що утворюютьсяепітелієм і сполучною тканиною, яка завжди під ним розташована та складається з аморфної речовини і волокон.
Залози зовнішньої, внутрішньої та змішаної секреції.
Залози змішаної секреції – залози що виконують зовнішню та внутрішню секреторну функції, в людей це підшлункова та статеві залози.
Основні ендокринні залози: 1 Шишкоподібне тіло 2 гіпофіз 3 Щитоподібна залоза 4 Тимус 5 Надниркова залоза 6Підшлункова залоза 7 Яєчники (жінки) 8 Яєчки(чоловіки) |
Підшлункова залоза бере участь у процесі травлення, тобто працює як залоза зовнішньої секреції, а з іншого виробляє інсулін таглюкагон - гормони, що регулюють вміст цукру у крові, тобто виконує внутрішню функцію.
Статеві залози – у жінок яєчники, у чоловіків - сім’яники або яєчка. Виконують як зовнішньосекреторну функцію продукуючи статеві клітини так і внутрішньосекреторну виділяючи у кров статеві гормони.
За́лози вну́трішньої секре́ції, також Ендокринні залози — залози, які не мають вивідних проток, і виділяють свою «продукцію» — гормони, безпосередньо в кров, яка омиває ці залози. Саме через це їх називають залозами внутрішньої секреції (ендокринні залози). У людини до ендокринних залоз відносять: епіфіз,гіпофіз, щитоподібну, загрудинну, паращитоподібні, наднирники, острівці Лангерганса підшлункової залози та ендокринну частину статевих залоз. У зв'язку з цим підшлункову залозу та статеві залози відносять до змішаних залоз за типом секреції.
Ендокринні залози не мають вивідних протоків, секрет виділяється в кров, а з неї надходить у міжклітинну рідину. Основна тканина — залозистий епітелій. Залози добре кровопостачаються та інервуються.
Ендокринні залози функціонують не ізольовано, а системно. Більшість гормональних процесів регулюється гіпофізом, який виділяє тропні гормони, що регулюють діяльність інших залоз. В свою чергу, секреторну діяльність гіпофіза регулює гіпоталамус, нейрони якого продукують нейроендокринні трансмітери, що стимулюють або пригнічують роботу гіпофіза. Ці два органи утворюють єдину гіпоталамо-гіпофізарну систему, в якій перший виконує регулюючу роль, а інший — ефекторну.
Наука, щи вивчає залози внутрішньої секреції — ендокринологія.
Залоза зовнішньої секреції, або Екзокринна залоза (від грец. éxo — поза, зовні і грец. krínó — виділяю) — залоза, що виділяє через вивідні протоки речовини, які виробляє організм в результаті процесів життєдіяльності, на поверхню тіла або слизових оболонок, у ті чи інші порожнини. Розвинені у людей і тварин.
Класифікація екзокринних залоз.
Екзокринні залози (від екзо... і греч.(грецький) krínó — відокремлюю, виділяю), залози зовнішньої секреції, залози тварин і людини, що виділяють через вивідні протоки речовини, що виробляються ними, на поверхню тіла або слизистих оболонок, в ті або інші порожнини. До екзокринних відносяться сальні, слинні, потові, молочні, слізні, мускусні залози, а також печінка, залози шлунково-кишкового тракту і ін., що виділяють секрет в порожнину травного каналу.
Екзокринні залози відрізняються будовою, формою, механізмом секреції і характером секрету. За будовою ці залози поділяються на одно- і багатоклітинні. У хребетних тварин та людини одноклітинні залози звичайно розміщуються в товщі епітелію, а найбільш поширеною формою їх є бокалоподібні клітини (мають розширену верхівку та звужену основу), які виділяють слизовий секрет.
Більшість екзокринних залоз складається з багатьох клітин і має складну будову. Залежно від будови і форми вивідних протоків та кінцевих відділів вони бувають трубчасті, альвеолярні та трубчасто-альвеолярні. Вони можуть бути також простими й складними, розгалуженими і нерозгалуженими.
Існує ще класифікація залоз, яка враховує спосіб секреції та вид секрету. Залежно від виду секрету залози поділяються на слизові, серозні та серозно-слизові. За способом секреції розрізняють залози мерокринні (еккринні), апокринні та голокринні.
Утворення та виділення секрету в мерокринних залозах відбувається без порушення цілісності залозистих клітин. В апокринних залозах під час виділення секрету верхівка клітини разом з гранулою секрету відривається. У людини до таких залоз відносять молочні та деякі потові. Залози з голокринним типом секреції у хребетних тварин та людини трапляються рідко. До них відносять сальні залози. При виділенні секрету — сала (sebum) відбувається повне руйнування залозистих клітин.
Крім морфологічних та функціональних класифікацій існує ще поділ епітеліальної тканини за походженням на п'ять типів: шкірний, кишковий, нирковий, целомічний та епендимогліальний. Кожен тип епітелію виконує певну функцію і, відповідно до цього, має певну будову.
Регенерація в епітеліальній тканині.
Оскільки клітини поверхневого епітелію контактують із навколишнім середовищем, вони часто пошкоджуються і відмирають. Тому епітеліальній тканині властива висока здатність до відновлення (регенерації). Зокрема, здатність епітелію до регенерації є однією з умов загоєння ран.
Регенерація епітелію здійснюється за рахунок наявності камбіальних клітин, різних для кожного виду епітелію. З фармацевтичних препаратів, присутніх на ринку України, регенерацію епітелію шкіри прискорюють Левосин, Метилурацилова мазь, Вундехіл, Пантенол та ін. Існує дві класифікації епітеліальної тканини: філогенетична (або просто генетична) і морфофункціональна. Згідно з філогенетичною класифікацією, запропонованою М.Г. Хлопіним, розрізняють шкірний, кишковий, нирковий, целомічний, епендимогліальний та ангіодермальний епітелії. Більш вживаною класифікацією є морфофункціональна класифікація, в основі якої лежать особливості будови і функції різних видів епітелію.
Загальна характеристика сполучних тканин або тканин внутрішнього середовища.
Сполучні тканини об’єднують під загальною назвою групу тканин, які називаються так тому, що утримують на місці клітини інших тканин. Здатність сполучної тканини зв’язувати інші тканини, а також служити для них опорою зумовлена тим, що певні клітини сполучної тканини виділяють міжклітинний матеріал, у деяких випадках дуже щільний, котрий і відповідає за підтримку цілісності тіла та надання йому певної форми. Різні типи сполучної тканини виконують багато інших функцій, причому деякі з них майже не є справді зв’язувальними. Навіть власне сполучні тканини містять іноді клітини, які виконують функції іншого роду.
Тканини внутрішнього середовища - це велика група тканин, які разом з епітелієм належать до так званих загальних тканин. Тканинами внутрішнього середовища є кров, лімфа і сполучна тканина з усіма її різновидами. Незважаючи на те, що окремі різновиди тканин внутрішнього середовища за зовнішніми ознаками значно відрізняються між собою (наприклад, кров і кісткова тканина), є всі підстави для поєднання їх у єдиний тканинний тип, а саме: спільність походження, будови і функції.
Спільність походження цих тканин є найвагомішою ознакою і полягає у тому, що всі вони розвиваються з мезенхіми. Мезенхіма - найпримітивніша сполучна тканина, яка існує лише на ранніх стадіях ембріонального розвитку. За будовою мезенхіма нагадує сітку, тому що клітини її мають зірчасту або веретеноподібну форму і контактують одна з одною своїми відростками. У петлях сітчастого остова міститься драглиста маса - міжклітинна речовина, щільність якої коливається зі змінами обміну речовин. Із мезенхіми шляхом диференціації розвиваються кров, лімфа і всі види сполучної тканини.
Функції тканин внутрішнього середовища різноманітні, але їх звичайно поєднують під загальною назвою "опорно-трофічні тканини". Вони виконують такі функції: трофічну, захисну, опорну (механічну). Функціональні особливості різних видів тканин внутрішнього середовища значною мірою залежать від фізико-хімічних властивостей міжклітинної речовини.
Класифікація тканин внутрішнього середовища.
Спільність будови цих тканин полягає у наявності міжклітинної речовини, яка у кількісному відношенні переважає над клітинами. На основі будови міжклітинної речовини можна виділити основні типи тканин внутрішнього середовища: кров та лімфа; власне сполучна тканина; хрящова тканина; кісткова тканина.
До основних груп тканин, що об’єднують у категорію сполучних, належать:
1) власне сполучна тканина;
2) жирова тканина;
3) кров та кровотворні тканини;
4) хрящова тканина;
5) кісткова тканина.
Варто зазначити, що різні автори класифікують сполучні тканини по-різному, тому в деяких джерелах можна зустріти іншу класифікацію цієї тканини.
Власне сполучна тканина виконує дуже важливу роль, зв’язуючи різні компоненти тіла.
Власне сполучна тканина являє собою групу сполучних тканин зі спеціальними властивостями, а також пухку сполучну та щільну сполучну тканини. Щільна сполучна тканина майже цілком складається з міжклітинної речовини, яка містить у великій кількості колагенові волокна, дуже міцні і майже не здатні до розтягування, та клітин — фіброцитів, що їх продукують. Ця тканина дуже міцна, з неї побудовані сухожилля, апоневрози, зв’язки та глибокий сполучнотканинний шар шкіри. Пухка сполучна тканина менш спеціалізована і містить не тільки всі основні види міжклітинної речовини та клітини, що її утворюють (фібробласти), але й інші клітини, що виконують специфічні функції (макрофаги, тканинні базофіли, плазмоцити та ін.). Пухка сполучна тканина має вигляд тонкого прошарку між сусідніми органами, напр., «одягає» зовні кровоносні судини, нерви, м’язи і фасції. Крім опорно-механічної функції, пухка сполучна тканина виконує захисну і трофічну функції, а також бере участь у пластичних процесах при загоюванні ран, утворенні капсули навколо стороннього тіла тощо. Утворенню сполучнотканинних рубців запобігає фармацевтичний препарат Контрактубекс.
Жирова тканина, заповнюючи вільні простори у сполучній тканині і в багатьох ділянках тіла, виконує амортизаційну та терморегулювальну функції, здійснює запасання жиру. Жирова тканина складається більшою мірою з жирових клітин (адипоцитів, ліпоцитів), які накопичують жир, пігментоцитів (меланоцитів), які входять до складу шкіри і мають коричневий пігмент — меланін, а також міжклітинної речовини.
Хрящі, кістки та суглоби виконують зв’язувальну та опорну функції завдяки своїй міжклітинній речовині. Хрящова тканина являє собою спеціалізований різновид сполучної тканини, яку відносять до групи тканин з опорною та формовірною функцією. Основні компоненти хрящової тканини — це міжклітинний матрикс з великою кількістю колагенових та еластичних волокон та клітини (хондроцити), які продукують матрикс.
Хрящова тканина містить добре розвинену основну міжклітинну речовину та волокна, в результаті чого цей вид тканин внутрішнього середовища характеризується великою міцністю й пружністю і належить до так званих скелетних тканин.
Кісткова тканина за будовою та функціональним значенням є унікальним різновидом сполучної тканини, яка містить велику кількість мінеральних солей (найбільше сполук кальцію та фосфатів), що становить 50–70% сухої маси кістки. У зв’язку з цим при різних патологіях опорно-рухового апарату людини лікарі призначають препарати кальцію. Кісткова тканина містить добре розвинену основну міжклітинну речовину. Високий ступінь мінералізації (біля 70% кісткової тканини складають фосфорнокислі солі кальцію у формі кристалів гідроксиапатиту) та товсті паралельні пучки колагенових волокон (так звані кісткові пластинки) забезпечують міцність кістки.
Клітини крові та лімфи не виконують власне зв’язувальної ролі в організмі, вони відповідають за виконання специфічних функцій, тому деякі автори виділяють їх у п’яту групу тканин.
Кров, її склад.
Кров (sапgиіs) - це рідка тканина організму, що циркулює у системі замкнених трубок-судин. Кров становить 1/13, або 5-9 %, маси тіла, що у дорослої людини дорівнює приблизно 5,0-5,5 л. Кров складається із рідкої частини - плазми, яка займає 55-60 % об'єму, і формених елементів, об'єм яких 40-45%. Плазма - це міжклітинна речовина крові. До формених елементів крові належать еритроцити, лейкоцити та тромбоцити (кров'яні пластинки).
Кров виконує низку життєво важливих функцій. Захисна функція крові полягає у забезпеченні гуморального і клітинного імунітету. Дихальна функція забезпечується шляхом переносу кисню та вуглекислоти. Суть трофічної функції - перенесення поживних речовин. Екскреторна функція полягає у виведенні шлаків. Гуморальна функція забезпечується шляхом транспорту гормонів та інших біологічно активних речовин. Гомеостатична функція полягає у підтриманні сталості внутрішнього середовища організму, в тому числі імунного гомеостазу.
Плазма крові - це колоїдний розчин, в'язкість якого у 5 разів вища, ніж в'язкість води. Плазма містить у собі 90-93% води та 7-10% сухого залишку. В останньому близько 7% складають білки і 3% - інші органічні та мінеральні речовини. Загальна концентрація мінеральних речовин у плазмі крові становить 0,9%; рН плазми 7,36.
До білків плазми належать:
альбуміни, які становлять близько 4%; вони зв'язують та переносять з кров'ю цілу низку речовин;
глобуліни становлять 1,1-3,1%, поділяються на альфа-, бета- і гамма-глобуліни (імуноглобуліни); в останній фракції містяться антитіла;
фібриноген, кількість якого 0,2-0,4%, важливий тим, що завдяки його здатності переходити у нерозчинну форму - фібрин - здійснюється процес згортання крові.
Плазма, з якої видалений фібрин, називається сироваткою крові. Це жовтувата, прозора рідина, яка використовується для виготовлення багатьох лікарських препаратів.
Еритроцити. їх склад та функції.
Еритроци́ти — або червонокрівці, у ссавців і людини є нерухомими, високодиференційованими клітинами, які у процесі розвитку втратили ядро та всі цитоплазматичні органели і пристосовані до виконання практично єдиної функції - дихальної, що здійснюється завдяки наявності в них дихального пігменту гемоглобіну. Тобто еритроцити, це невеликі без'ядерні клітини крові червоного кольору, функція яких — транспорт кисню і вуглекислого газу.
Загальна кількість еритроцитів у крові однієї людини становить близько 25×1012. Загальний об'єм еритроцитів у людини - 2л. Під час аналізів крові вміст всіх формених елементів подається на одиницю об'єму - 1л.
Позначають еритроцити скороченням латинськими літерами Er. Кількість еритроцитів дорівнює у чоловіків від 3,9×1012 до 6,0×1012 в 1л. у жінок - від 3,7×1012 до 5,5×1012 в 1л. Більша концентрація еритроцитів спостерігається у крові новонароджених дітей - від 6,0×1012 до 9,0×1012 в 1л, а також у старих людей - до 6,0×1012 в 1л. Число еритроцитів у здорових людей може коливатися залежно від фізичного навантаження, перебування в умовах розрідженої атмосфери, дії гормонів тощо. Зокрема, жіночі статеві гормони гальмують розвиток еритроцитів, унаслідок чого вміст червонокрівців у крові жінок менший, ніж у чоловіків. Підвищення кількості еритроцитів в одиниці об'єму крові називається еритроцитоз або поліцитемія , а зниження — еритроцитопенія.
Еритроцити у людини і ссавців здебільшого мають форму двоввігнутих дисків, їх називають дискоцитами. У нормі дискоцити становлять 80% від загальної кількості еритроцитів. Трапляються й інші форми еритроцитів - планоцити ( мають плоску поверхню), сфероцити (кулясті), ехіноцити ( мають шипи) тощо. Така різноманітність форм у нормі позначається терміномфізіологічний пойкілоцитоз. Коли ж кількість змінених форм еритроцитів перевишує 20%, те ж явище має назву патологічного пойкілоцитозу. Форму еритроцитів підтримує бета-сіа-глікопротеїнв еритроцитарній мембрані та спеціальний каркас, побудований збілка спектрину, який прилягає до плазмолеми і пов'язаний з нею йншим білком - анкерином.
Діаметр еритроцита у людини 7,1 - 7,9 мкм, товщина клітини на краях 2-2,5 мкм, у центрі - до 1мкм. Заглибина еритроцита в тонкій центральній частині має назву фізіологічної ескавації. Така форма клітини забезпечує збільшення її поверхні і прискорює насичення гемоглобіну киснем. В умовах норми 75% усіх еритроцитів мають вищеназвані розміри. Це так звані нормоцити. Частина клітин має діаметр понад 8 мкм Це макроцити, їх кількість - 12,5%. Решта еритроцитів може мати діаметр 6 мкм і менший. Це мікроцити. Якщо кількість макро- і мікроцитів перевищує 25%, це явище має назву анізоцитозу.
Під світловим мікроскопом у мазках крові еритроцити мають вигляд безструктурних округлих дисків, фарбуються оксифільно. Оксифілія зумовлена наявністю гемоглобіну. Центральна (тонка) частина еритроцита фарбується менш інтенсивно. Електронна мікроскопія свідчить, що еритроцит покритий плазмолемою товщиною близько 20 нм. На її зовнішній поверхні розташовані антигенні олігосахариди, які зумовлюють групову належність еритроцитів, фосфоліпіди, сіалова кислота. Усередині еритроцитів розташований електронно-щільний вміст - численні гранули гемоглобіну розмірами 4-5 нм.
За хімічним складом еритроцити мають 60% води і 40% сухого залишку.95% сухого залишку складає гемоглобін і лише 5% - інші речовини. Таким чином, гемоглобін становить одну третину загальної маси еритроцитів. У крові дорослої людини міститься близько 600г гемоглобіну, тобто в 100г крові 15г гемоглобіну. Гемоглобін є пігментом, який надає крові червоного кольору.
Гемоглобін зв'язуючи кисень переносить його і віддає периферійним тканинам. Гемоглобін, який віддав кисень називається відновленим або редукованим, він має колір венозної крові. Віддавши кисень кров поступово вбирає в себе продукт обміну речовин СО2 (вуглекислий газ). Гемоглобін який зв'язує вуглекислий газ називається карбогемоглобіном.
Скорочення кількості гемоглобіну у еритроцитів крові називається анемією.
Основна функція еритроцитів – перенесення кисню від легень до тканин і вуглекислого газу від тканин до легень. Газообмінорганізму з навколишнім середовищем здійснюється таким чином. Проходячи через капіляри легень, гемоглобін легко приєднує кисень і перетворюється у нетривку сполуку — оксигемоглобін, що у тканинах інших органів розщеплюється виділяючи при цьому кисень, який використовується клітинами тканин. Звільнений від кисню гемоглобін відразу приєднує вуглекислий газ – продукт розпаду речовин у клітинах. Також еритроцити забезпечують підтримку рН крові (гемоглобін і оксигемоглобін складають одну з буферних систем крові); підтримання іонного гомеостазу за рахунок обміну іонами між плазмою та еритроцитами; участь у водному і сольовому обміні; адсорбція токсинів, в тому числі продуктів розпаду білка, що зменшує їх концентрацію в плазмі крові і перешкоджає переходу в тканині; участь у ферментативних процесах, у транспорті поживних речовин - глюкози, амінокислот.
В кожному міліметрі кубічному крові міститься близько п'яти мільйонів еритроцитів, і майже 25000 мільярдів (25x1012) в організмі в цілому.
Види лейкоцитів.
Лейкоцити більше еритроцитів і містяться в крові в набагато меншій кількості (приблизно 7000 в 1 мм3 крові). Вони грають важливу роль в захисті організму від хвороб. Кожен лейкоцит має ядро. Незважаючи на наявність ядра, тривалість їхнього життя вкровотоці зазвичай не перевищує кількох днів. Всі вони здатні до амебоідному руху. Це дозволяє їм протискуватися через стінки капілярів в області контакту клітин ендотелію і направлятися до інфікованих тканин.
Лейкоцити можна бачити за допомогою світлового мікроскопа тільки в тому випадку, якщо вони пофарбовані. На забарвлених препаратах чітко виявляються дві основні групи лейкоцитів - гранулоцити, або зернисті лейкоцити,містять в цитоплазмі гранули, і агрануло-ціти, або незерністие лейкоцити, які не мають таких гранул.