

ГОРЮНЫ Занятие I
Гормоны это органические биологически активные вещества, различные по химической природе, вырабатываемые в специализированных органах и тканях (железах внутренней секреции), поступающие непосредственно в кровь и осуществляющие гуморально регуляцию обмена веществ и функций организма.
Гормоноидами называют вещества, вырабатываемые в ряде тканей и клеток (не в специализированных органах), подобно гормонам влияющие на обменные процессы и функции организма. Гормоноиды часто оказывают свое действие внутри тех клеток, в которых они образуются, или же они распространяются путем диффузии и действуют вблизи места своего образования, некоторые же гормоноиды попадают и в кровоток. Резких различий между гормонами и гормоноидами нет.
Гормоны эндокринной системы совместно с нервной системой осуществляют интеграцию обменных процессов, протекающих в клетках, тканях, органах, в интересах целого организма.
Железами внутренней секреции являются гипофиз, эпифиз, щитовидная 'железа, паращитоввдные железы, поджелудочная железа (островковый аппарат), надпочечники, половые железы, тимус, кроме того, гормоны продуцируются гипоталамусом, плацентой, стенкой пищеварительного тракта, почками, печенью, сердцем, тучными клетками соединительной ткани, а простагландины образуются почти во всех клетках. Клетки органов и тканей, способные трансформировать аминокислоты в различные гормоны (в том числе и пептидной природы), называют апудоцитами, а их совокупность - АПУД-системой организма. Апудоциты обнаруживают практически во всех органах.
Синтезированные гормоны депонируются в соответствующих железах в разных количествах: запас стероидных гормонов невелик - его хватает на обеспечение организма лишь в течение нескольких часов, запаса белково-пептидных гормонов (в форме прогормонов) хватает на одни сутки, катехоламины накапливаются в количестве, обеспечивающем организм в течение нескольких суток, запаса же тиреоиднкх гормонов хватает на несколько недель. Секреция гормонов в кровь (путем экзоцитоза или диффузии) происходит неравномерно - она носит пульсирующий характер, или же наблюдается циркадный'ритм. В крови белкоро-пептидщш гормоны

 и
катехоламины
обычно
находятся
в
свободном
состоянии,
стероидные
же
и
тиреоидине
гормоны
связываются
со
специфическими
белками-переносчл-ками.
Период
полужизни
гормонов
в
плазме
составляет:
катеходаминов
-секунды,
белково-пептидннх
гормонов
- минуты,
стероидных
гормонов
-часы,
тиреоидных
гормонов
- несколько
суток.
Гормоны
воздействуют
на
клетки-мишени,
взаимодействуя
с
рецепторами,
их
отделение
от
рецедто-ров
происходит
через
десятки
секунд
или
минуты.
Все
гормоны
в
конечном
счете
метаболизируются
(разрушаются),
частично
в
клетках-мишенях,
особенно
же
интенсивно
- в
печени.
Выделяются
из
организма
главным
образом
метаболиты
гормонов,
неизмененные
гормоны
- лишь
в
очень
малых
количествах.
Основной
путь
их
выведения
- через
почки
с
мочой.
и
катехоламины
обычно
находятся
в
свободном
состоянии,
стероидные
же
и
тиреоидине
гормоны
связываются
со
специфическими
белками-переносчл-ками.
Период
полужизни
гормонов
в
плазме
составляет:
катеходаминов
-секунды,
белково-пептидннх
гормонов
- минуты,
стероидных
гормонов
-часы,
тиреоидных
гормонов
- несколько
суток.
Гормоны
воздействуют
на
клетки-мишени,
взаимодействуя
с
рецепторами,
их
отделение
от
рецедто-ров
происходит
через
десятки
секунд
или
минуты.
Все
гормоны
в
конечном
счете
метаболизируются
(разрушаются),
частично
в
клетках-мишенях,
особенно
же
интенсивно
- в
печени.
Выделяются
из
организма
главным
образом
метаболиты
гормонов,
неизмененные
гормоны
- лишь
в
очень
малых
количествах.
Основной
путь
их
выведения
- через
почки
с
мочой.
2. По своей хитагаеской природе гормоны могут быть белками (простыми или сложными - гликопротеинами), пептидами, стероидами или же они являются производными аминокислот. Простагландины, простациклин, тромбоксаны и лейкотриены возникают из полиненасыщенных жирных кислот.
Гормонами белковой природы являются инсулин, паратирин, гормоны передней доли гипофиза (кроме кортикотрошна, эндорфинов, энкефалинов и когерина), а также плацентарные (хорионические) гормоны, аналогичные гипофизарным, я релаксин.
К гормонам-пептидам относятся кортикотропин, эндорфины, энкефа-лины, когерин, меланотрогшн, вазопрессин и окситоцин гипофиза, либе-рины и статины гипоталамуса, кальцитонин, глюкагон, большинство гормонов тимуса, гормоны пищеварительного тракта.
Стероидными гормонами являются гормоны коры надпочечников, а также мужские и женские половые гормоны.
К производным аминокислот относятся катехоламины (возникают из тирозина), тиреоидине гормоны (также образуются из тирозина), мелато-нин и серотонин (возникают из триптофана).
1 &i Тироксин синтезируется в щитовидной железе, 11-ДОК - в коре надпочечников, дофамин возникает в мозговом слое надпочечников, в центральной нервной системе.
3,5,3^-Трийодтаронин синтезируется в щитовидной железе, эст-радиол - в яичниках и плаценте (в некоторых количествах обнаруживается и в семенниках, и в коре надпочечников), серотонин в наибольших количествах возникает в клетках кишечника, э также в центральной нервной системе, в эпифизе).

5ес-Дигвдротейтостерон возникает в семенниках и в очень небольших количествах в клетках-мишенях из тестостерона, кортизол образуется в коре надпочечников, гистамина особенно много в тучных клетках соединительной ткани, много его образуется в слизистой желудка, возникает он в головном мозгу и в других тканях.
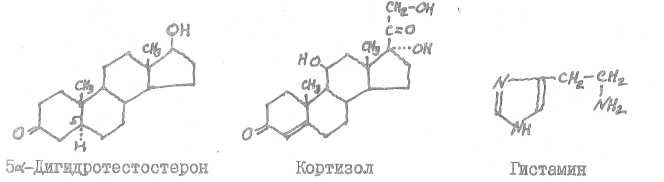
Кортикостерон синтезируется в коре надпочечников, эстриол возникает в яичниках, в плаценте, ацетилхолин является медиатором холив-ергическш; систем, он образуется в нервно-мышечных синапсах, в парасимпатической нервной системе.
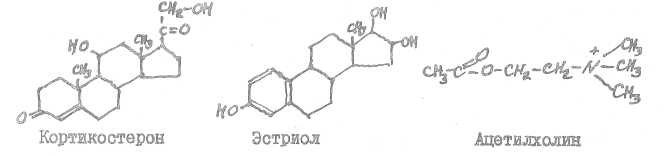
Норадреналин образуется в 'Центральной нервно! системе, в мозговом слое надпочечников 9 альдостерон - в коре надегочечнщюв8 эстрон (фолликулин) возникает в яичниках, в плаценте,,
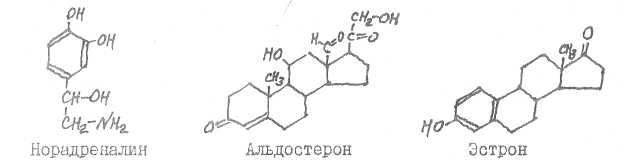
Мелатошш синтезируется в эпифизе, кортизон - в коре надпочечников, андростерон возникает в печени в процессе метаболизма тестостерона.
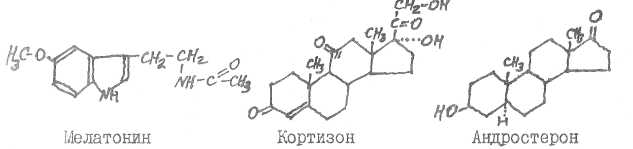
Гидрокортизон (кортизол) образуется в коре надпочечников, тестостерон - в семенниках (некоторое количество его возникает и в плаценте, в яичниках, в коре надпочечников), 17-кетостероиды образуются в печени.
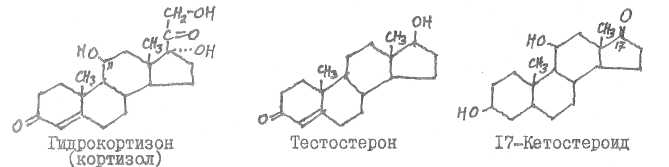
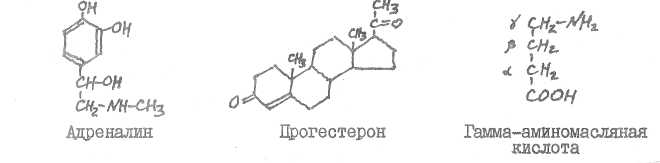
Адреналин образуется в мозговом веществе надпочечников, в симпатической нервной системе, в центральной нервной системе, прогестерон синтезируется в желтом теле, в плаценте, гамма-аминомасляная кислота - в центральной нервной системе.
Гормоны являются регуляторами обменных процессов и функций организма. Основными биохимическими эффектами действия гормонов на
клетки-мишени являются изменение проницаемости плазматической мембраны, изменение активности различных ферментов, изменение концентрации ферментых молекул в клетке. Всё это приводит к изменениею функции клетки, ткани, органа.
12. Белковые и пептидные гормоны, а также и катехолаиияы действуют на рецепторы наружной поверхности плазматической мембраны. Это воздействие приводит к активированию (или ингибированию) мембранной аденилатциклазы (гуанилатциклазы) с появлением вторичного посредника гормонального действия (цАМФ или ц1Ш), активированию в цитоплазме различных протеинкиназ, которые в свою очередь фосфорилируют различные клеточные белки (в том числе и ферменты) цитоплазмы, мембран, митохондрий, ядра, рибосом. В результате изменяются каталитическая активность ферментов, интенсивность биосинтеза белков, проницаемость мембран, а, следовательно, изменяется и функционирование клеток.
Воздействие некоторых белково-пелтидных гормонов и катехолами-нов на плазматическую мембрану клетки может вести к усиленному поступлению в цитоплазму ионов Са^+ (вторичный посредник) извне через кальциевые каналы или из митохондрий (под влиянием цАМФ), или же ионы Са высвобождаются из связи с фосфорилированяыми молекулами фосфа-тидилинозитов плазматический мембраны. Ионы Са^+ взаимодействуют с белком кальмодулином, а возникающий комплекс кальций-кальмодулин активирует протеинкиназы, в дальнейшем фосфоршшрующие различные клеточные белки, что и ведет к изменению функции клетки.
Роль вторичного посредника в действии ряда гормонов на клетки-мишени выполняет также 2^адеяиловый олигонуклеотид (2^!>-олигоадени-лат или олиго(А) из 3-4 мономеров). Он образуется в клетке под влиянием цАМФ и активирует особые рибояуклеазы, гидролизующие клеточные мРНК и рРНК, вследствие чего тормозится синтез определенных белков, замедляется клеточное деление (антилролиферативный эффект).
Образование в клетках и действие вторичных посредников (мессвнд-жеров) гормонального действия цАМФ, ионов Са и олиго(А) взаимосвязаны. Так, увеличение содержания цАМФ приводит к выбросу Са2+ из митохондрий в цитоплазму и к росту концентрации олиго(А), в то же время Са + и олиго(А), активируя фосфодиэстеразу, вызывают снижение уров-ня цАМФ. Влияние указанных посредников гормонального действия в клетке может быть направлено на одни и те же биохимические механизмы, которые в этом случае находятся год контролем сразу двух (или нескольких) вторичных мессанджеров, например, цАМФ и Са2+, цАИ и олиго(А) и т.д. Поэтому цАМФ, олиго(А) и Са по-видимому являются в клетках
к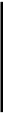
 омпонентами
единой
регуляторной
системы.
омпонентами
единой
регуляторной
системы.
Роль вторичных посредников гормонального действия в клетках-мишенях выполняют также особые пептиды, диацилглицерол, инозитолгри-фосфат, арахвдоновая кислота, а также простагландины и другие еще неизученные вещества. Последние данные показывают, что многие пептидные гормоны с поверхностным мембранным рецептором образуют комплекс гормон-рецептор, который быстро перемещается внутрь клетки, причем рецептор в дальнейшем проникает в ядро и воздействует на генетический аппарат. В этих случаях сам рецептор выполняет роль вторичного посредника гормонального действия.
Прекращение действия гормонов на клетки достигается отделением гормона от рецептора, гидролизом ГТФ У-белком, гидролизом циклического нуклеотвда фосфодиэстеразой, а также действием фосфопротеинфое-фатазы.
Активирование гормонами ряда ферментов (например, гликогенфоо-форилазы, гликогенсинтазы) имеет многоступенчатый, каскадный характер. Биологический смысл этого заключается в том8 что на каждом этапе происходит многократное усиление первичного гормонального сигнала, и весь процесс активирования ферментов носит взрывной характер. Так, одна молекула гормона обеспечивает появление около 500 молекул цАМФ„ каждая из которых, в свою очередь» активирует множество молекул про-теинкиназы и т.д.
13. Стероидные гормоны проникают в ядро клеток-^шшеней9 где сое диняются с рецепторами. Комплексы гормон-рецептор взаимодействуют с негистоновыми белками хроматина, в результате становятся доступными
. для ШК-полимеразы соответствующие участки ДНК. Тем самым облегчается синтез мШК, стимулируется транскрипция, усиливается синтез белков. Тиреоидине гормоны взаимодействуют с рецепторами' в хроматине ядра клеток-мишеней, усиливают синтез мШК и, следовательно, синтез белков, втом числе многих ферментов. Возможно, тиреоидине гормоны влияют и на механизм трансляции.
14. Циклические нуклеотвды (цАМФ и цГМФ) выполняют роль вторич ного посредника в действии на клетки-мишени гормонов, взаимодействую щих с рецепторами плазматической мембраны (белково-пептидные гормоны, катехоламины).
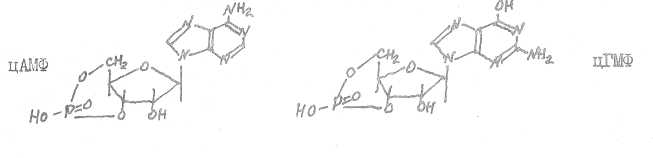
Мембранный фермент аденилатциклаза катализирует образование из АТФ в цитоплазме клетки-адишени циклического АМФ, являющегося вторичным посредником в действии белковых, пептидных гормонов и ка-техоламинов.
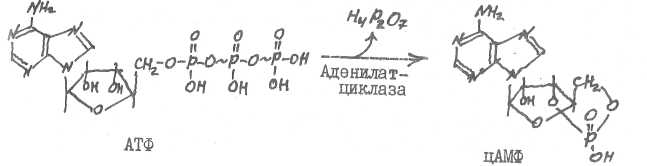
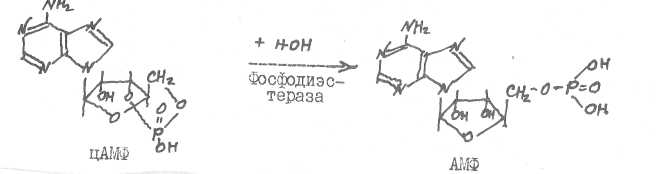
Фосфодиэстераза гидролизует цАМФ (цШФ), прерывая тем самым процесс воздействия гормонов (белковых, пептидных, катехоламинов) на клетки-мишени.
В щитовидной железе вырабатываются гормоны: тироксин или 3,5,3',5- тетрайодтиронин (обозначается сокращенно Т4> он представляет собой тирозин, соединенный с гидроксифенилом и четырежды йодированный), 3,5,3-трийодтирония (Тд) и кальцитонин (пептид из 32 аминокислот) .
Йодсодержащие тиреоидине гормоны повышают основной обмен организма, ускоряют рост и дифференцировку тканей. Они усиливают поглощение кислорода и теплопродукцию большинством тканей, интенсифицируя обмен углеводов, липидов, белков, повышают активность более 100 различных ферментов. Термогенное действие этих гормонов в основном обусловлено их стимулирующим влиянием на мембранную Л'а+,К+-АТФазу, функционирование которой, связанное с гидролизом значительного количества АТФ, является одним из основных источников тепла в организме. Ти-реовдные гормоны также активируют матохондриальную глицерол-3-фосфат-дегидрогеназу, усиливая тэм самым функционирование глицерофосфатного челночного механизма транспорта водорода из цитоплазмы в митохондрии, что ведет к сокращению синтеза АТФ и увеличению продукции тепла (по сравнению с эффектом, производимым малатным челночным механизмом).
В
 печени
тиреовдяые
гормоны
стимулируют
окисление
внемитохондриально-го
НАДН(Н+)
с
помощью
цитохрома
#5,
локализованного
во
внешней
мембране
митохондрий
и
передающего
водород
на
цитохром
с,
что
обеспечивает
значительную
теплопродукцию.
Тиреоидине
гормоны
также
стимулируют
синтез
/S-адренергических
рецепторов,
усиливая
действие
на
клетки
катехоламинов.
печени
тиреовдяые
гормоны
стимулируют
окисление
внемитохондриально-го
НАДН(Н+)
с
помощью
цитохрома
#5,
локализованного
во
внешней
мембране
митохондрий
и
передающего
водород
на
цитохром
с,
что
обеспечивает
значительную
теплопродукцию.
Тиреоидине
гормоны
также
стимулируют
синтез
/S-адренергических
рецепторов,
усиливая
действие
на
клетки
катехоламинов.
Т3 обладает наибольшей биологической активностью, большая его часть возникает в периферических тканях путем дейодирования Т^.
Кальцитонин сникает уровень кальция и фосфора в крови, усиливая отложение фосфорно-кальциевых солей в костной матрице и в то же время тормозя реабсорбцию кальция и фосфора в почках.
Инактивирование йодсодержащих тиреоидных гормонов в периферических тканях (особенно в печени и почках) происходит путем их дейодирования, дезаминирования, декарбоксилирования с образованием в различной степени иодированных тиропирувата или тироацетата. Эти продукты могут подвергаться метилированию или образовывать глюкурониды. Кальцитонин метаболизируется путем гидролиза его пептидных связей.
I?. Сначала из пищевых иодидов под влиянием иодидперокоидааы и при участии Н^Оо возникает активный йод:
![]()
Тирозиниодиназа катализирует иодирование связанного в иодтирео-глобулине тирозина, затем протекает реакция конденсации с образованием связанного Т^ или Т^:
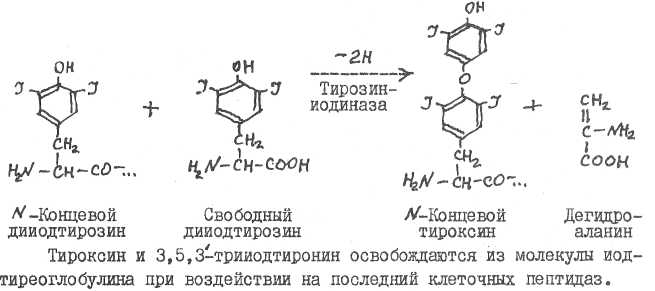
18. Основными видами патологии щитовидной железы являются базедова болезнь, кретинизм, микседема, эндемический зоб.
При базедовой болезни имеет место гиперпродукция тиреоидных гор-
монов, возникают симптомы, характерные для тиреотоксикоза - увеличение щитовидной железы (зоб), экзофтальм, усиленный основной обмен, повышенная температура тела, отрицательный азотистый баланс, гипергликемия, гилохолестеринемия, потеря веса, тахикардия и др.
Кретинизм представляет собой врожденную патологию щитовидной железы, сопровождающуюся нарушением синтеза тиреоидных гормонов, фи этом наблюдаются задержка роста, недостаточное физическое развитие, сильная умственная отсталость, сниженные основной обмен и температура тела.
Микседема (гипофункция щитовидной железы) проявляется снижением основного обмена и температуры тела, слизистым отеком, нарушением водно-солевого и липидного обмена, могут быть психические расстройства.
Эндемический зоб является следствием недостаточного поступления в организм йода с пищевыми продуктами и водой. При атом продукция тиреоидных гормонов либо не нарушена, либо снижена.
19. Паратгормон (паратирин) является белком (84 аминокислотных остатка). Он повышает концентрацию кальция в сыворотке крови за счет стимулирования остеокластического разрушения костного материала, а также усиления реабсорбции кальция в почечных канальцах и (косвенно) в кишечнике. Эти эффекты даратгормона обусловлены стимулированием им образования в почках 1,25-дигидроксихолекальциферола. Паратирин однако понижает концентрацию фосфора в крови благодаря торможению реабсорбции фосфатов в почках.
Недостаточность ларащитовидяых желез ведет к гипокальциемии, ги-перфосфатемии, а также к тетаническим судорогам.
20. Гормонами поджелудочной железы являются: инсулин (белок, содержит 51 аминокислотный остаток), глюкагон (пептид из 29 аминокислот), соматостатин (пептид из 14 аминокислот) и панкреатический полипептид (из 36 аминокислот).
Инсулин влияет на углеводный обмен (усиливает проникновение глюкозы из крови в клетки и ее утилизацию), на белковый обмен (анаболическое действие, стимулирует синтез белков), а также на обмен липи-дов (усиление липогенеза).
Глюкагон стимулирует катаболические процессы - мобилизацию гликогена в печени, глюконеогенез, гидролиз жиров в печени и жировой ткани.
Соматостатин ингибирует выделение поджелудочной железой инсулина, глюкагога и панкреатического полипедтида, а также тормозит секре-
цию гастрина, секретина.
Панкреатический полипептид влияет на функционирование желудочно-кишечного тракта.
21. Продукт трансляции инсулиновой мЕНК представляет собой пре-проинсулин (109 аминокислотных остатков), из которого возникают про-инсулин (86 аминокислот) и инсулин (51 аминокислота), поступающий в кровоток.
Инсулин стимулирует поступление глюкозы из крови в клетки (особенно мышц и жировой ткани), благодаря чему концентрация глюкозы в крови снижается, возникает гипогликемия. В клетках глюкоза фосфорили~ руется и интенсивно используется в качестве источника энергии путем ее катаболизма, чему способствует активирование инсулином гексокина-зы (глюкокиназы), Фосфофруктокиназы, пируваткиназы. Одновременно усиливается и синтез гликогена в печени (активируется гликогенсинтаза, ингибируется гликогенфосфорилаза). В то же время глюконеогенез тормозится за счет снижения под влиянием инсулина активности ключевых ферментов этого процесса.
Действует инсулин на клетки-мишени по-разному. Во-первых, взаимодействие молекул инсулина с рецепторами плазматической мембраны стимулирует тирозинкиназную активность рецептора, который начинает фосфорилировать белки мембраны ,в результате чего усиливается функционирование мембранных транспортных систем или эти системы переходят в функционально-активное состояние. Во-вторых, воздействие инсулина на мембранные рецепторы по-видимому вызывает высвобождение из мембранных структур небольшого пептида (возможно также, что этот пептид является частью молекулы самого инсулина), выполняющего роль вторичного посредника. Этот пептид ингибирует цАМФ-зависимую протеинкиназу, что ведет к изменению степени фосфоршшрованности клеточных белков, в том числе многих ферментов. Наконец, воздействие инсулина ведет к увеличению в клетке концентрации и такого вторичного посредника, как ионы Са2+. В результате происходят сдвиги в обмене углеводов, липи-дов, белков, нуклеиновых кислот. Инсулин также стимулирует фосфодиэс-теразу. Он усиливает синтез белков, жиров, нуклеиновых кислот. Механизмы этих влияний еще неясны.
Период полураспада инсулина в организме составляет 3-10 минут. Инсулин разрушается в основном в печени и почках. При этом сначала фермент глутатион-инсулин-трансгидрогеназа восстанавливает дисульфид-ные связи, вследствие чего молекула инсулина распадается на два полипептида, которые затем расщепляются инсулиназой.
Инсулиновая недостаточность в организме обусловливает развитие сахарного диабета. Она может возникнуть вследствие нарушения превращения проинсулина в инсулин, нарушения молекулярной структуры инсулина или его рецепторов, избыточного образования в организме диабето-генннх метаболитов триптофана, а также благодаря другим еще неизвестным причинам.
Сахарный диабет характеризуется гипергликемией, полиурией, глю-гозурией, полидипсией, кетонемией и яетонурией, высокой плотностью мочи, запахом ацетона от больного, истощением запасов гликогена в печени, усиленным распадом жиров и белков, частым исхуданием. Возможно развитие диабетической комы (кетоацидотической, гиперосмолярной или лактатацвдотической).
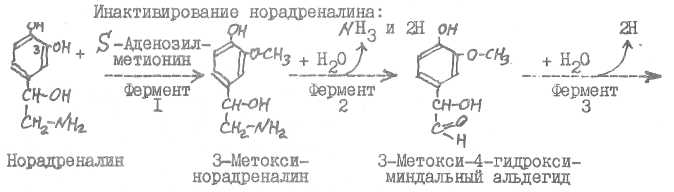
22. К катехоламинам относятся адреналин, норадреналин, дофамин. Термин "катехоламины" означает, что эти вещества в своей структуре содержат пирокатехиновое ядро и аминогруппу.
Возникают катехоламины из тирозина.
Основной путь инактивирования катехоламинов происходит путем метилирования их гидроксила в положении 3 бензольного кольца, дезамини-рования и окисления боковой цепи с образованием З-метокси-4-гидрокси-мин^льной кислоты (главный конечный продукт). Могут возникать и другие продукты инактявирования: 3-метокси-4~гидроксифенилгликоль, ванилиновая кислота, Л/-ацетилыше производные, глюкурониды, продукты хи-ноидного окисления (адренохром, адренолютив и др.).
Катехоламины влияют на углеводный и липидный обмены (норадреналин значительно слабее адреналина). Они усиливают распад гликогена в мышцах и мобилизацию гликогена в печени (возникает гипергликемия), они также стимулируют липолиз в жировой ткани и поступление жирных кислот в кровь. Дофамин выполняет функции медиатора в нервной системе.
Адреналин называют "гормоном тревоги", "аварийным" гормоном, так как он в трудных для организма условиях наиболее быстро обеспечивает мобилизацию энергетических (углеводных и жировых) ресурсов, одновременно стимулируя деятельность центральной нервной системы.
23» 24, 25 и 26 - см. стр. 96, 97.
27. Феохромоцитома - опухоль мозгового вещества надпочечников, продуцирующая избыточное количество катехоламинов. Проявляется она пароксизмами чувства страха, гипертонииs тахикардии, появляется гипергликемия, в крови повышается концентрация жирных кислот, содержание катехоламинов может возрастать в сотни раз, в моче резко увеличено
к оличество
З-метокси-4-гидроксиминдальной
кислоты.
Образование
дофамина
из
фенилалаяина:
оличество
З-метокси-4-гидроксиминдальной
кислоты.
Образование
дофамина
из
фенилалаяина:

Фермент I - фенилаланин-4-монооксигеназа (кофермент тетрагидро-биоптерин), фермент 2 - тирозин-3-моноокоигеназа (кофермент тетрагид-робиоптерин), фермент 3 - декарбоксилаза ароматических аминокислот. Ферменты I и 2 относятся к классу оксидоредуктаз, фермент 3 - к классу лиаз.
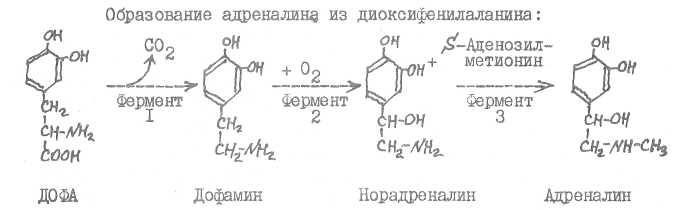
Фермент I - декарбоксилаза ароматических аминокислот, фермент 2 -дофаминмонобксигеназа (кофермент - аскорбиновая кислота), фермент 3 -норадрекалин-ААметилтрансфераза. Фермент I относится к классу лиаз, фермент 2 - к классу оксидоредуктаз, фермент 3 - к классу трансфераз.
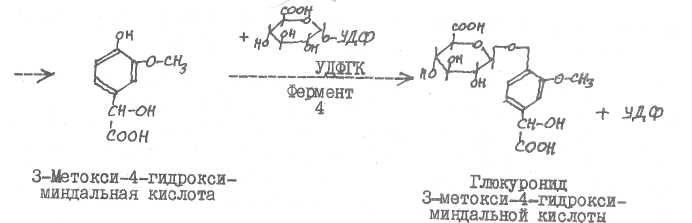
Фермент I - катехол-0-метилтрансфераза, фермент 2 - аминоксвда-за (ФМН.См), фермент 3 -альдегидоксидаза (ФАЦ. Мо), фермент 4 - УДФ-глюкуронозилтрансфераза. Ферменты I и 4 относятся к классу трансфераз, ферменты 2 и 3 - к классу оксидоредуктаз.
З-Метокси-4-гидроксиминдальная кислота - главный конечный продукт метаболизма катехоламинов.
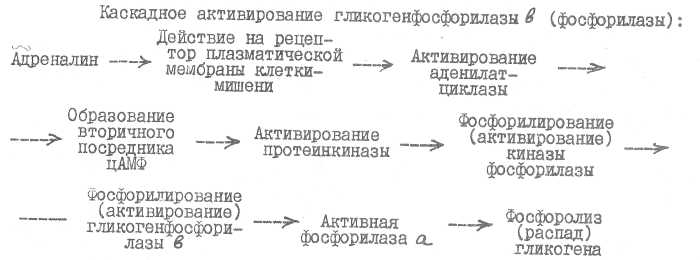
Распад гликогена приводит как к появлению свободной глюкозы в печени и гипергликемии, так и к интенсивному катаболизму глюкозы.с освобождением энергии.
Воздействие адреналина на гликогенсинтазу отличается тем, что активная протеинкиназа активирует путем фосфорилирования киназу гли-когенсинтазы, которая в свою очередь фосфорилирует активную гликоген» синтазу, вследствие чего гликогенсштаза инактивируется и синтез гликогена тормозится.
Феохромоцитома - опухоль мозгового вещества надпочечников, продуцирующая избыточное количество катехоламинов. Проявляется она пароксизмами чувства страха, гипертонии, тахикардии,.появляется гипергликемия, в крови повышается концентрация жирных кислот, содержа-
7-258 •
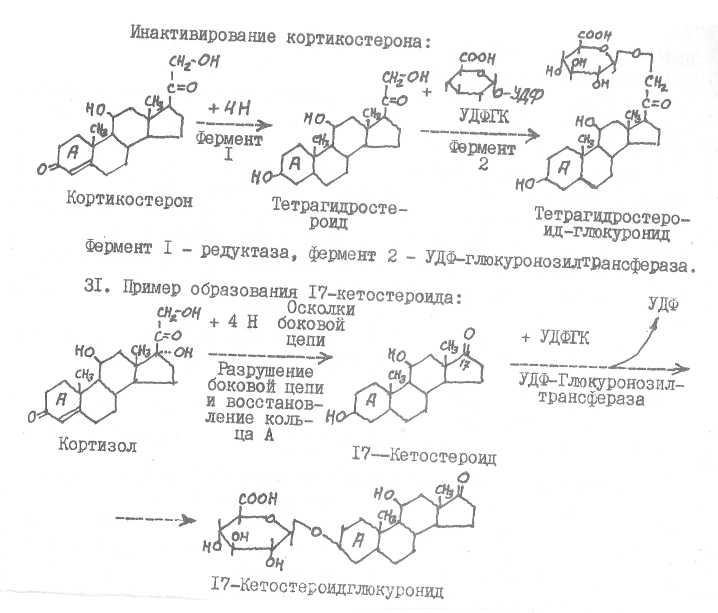
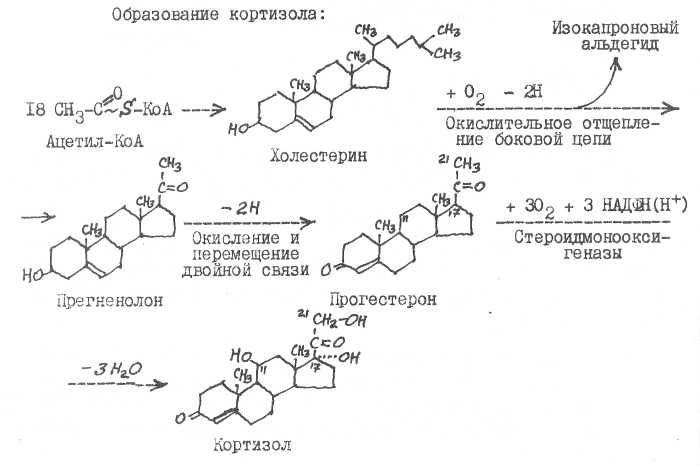
28. Истинными гормонами коры надпочечников являются кортизол (гидрокортизон) , кортикостерон, альдостерон. Высокой гормональной активностью обладают также кортизон и П-дезоксикортикостерон. Все эти кор-тикостероиды делят на глюкокортикоида (кортизол, кортизон, кортикостерон) и минералокортикоиды (альдостерон, II-ДОК, кортикостерон). В основе структуры гормонов коры надпочечников лежит ядро стерана.
Возникают кортикостероиды из холестерина.