

Пиримидиновые нуклеотиды
Теперь разберёмся в синтезе нуклеотидов и начнём с пиримидиновых. Но для начала представители. В основе пиримидиновых оснований лежит пиримидин, ему подобны урацил, тимин и цитозин (выучите их хорошо). Также вы должны знать источники атомов для пиримидина – они подписаны
в верхней части рисунка (см. рисунок ниже, углерод от углекислого газа, азот от аммиака, который в свою очередь предоставлен глутамином, остальное от аспартата). Первая реакция вам уже знакома из темы аминокислотного обмена – синтез карбомаилфосфата – аммиак, углекислый газ и ортофосфат от одной АТФ объединяются в единую молекулу за счёт энергии второй АТФ (одна АТФ – донор, другая – энергетик), фермент – карбомаилсинтетаза-2 (в отличии от версии 1 она расположена не в митохондриях, а в цитоплазме и имеет особую аллостерическую регуляцию: ФРПФ активирует, пиримидиновые нуклеотиды ингибируют.
121

Во второй реакции работает фермент карбомаиласпартаттрансфераза – от карбомаилфосфата отщепляется ортофосфати всё то, что осталось, сшивается, получается оротат – нарисовал в двух наклонах (слева для понимания, что откуда, а второй – как правильно пишется). Оротат – уже азотистое основание, пора делать нуклеотиды – третья реакция. Фермент оротатфосфорибозилтрансфераза берёт ФРПФ, отщепляет пирофосфат, а на его место ставит оротат – получаем оротидинмонофосфат ОМФ (нуклеотид), однако его карбоксильная группа нам ну никак не нужна – от неё избавимся
в четвёртой реакции. Фермент ОМФдекарбоксилаза отщепляет карбоксильную группу, тогда оротат превращается в урацил, а ОМФ в УМФ. А дальше последовательность реакций, в которой вы можете получить любой пиримидиновый нуклеотид. С помощью АТФ даёте ортофосфат УМФ,
122

превращая его в УДФ, дальше, если ходите дезоксирибонуклеотиды, подействуйте рибонуклеотидредуктазой, тогда УДФ превратится в дУДФ (рибоза превращается в дезоксирибозу), после этого вы можете взять аммиак от глутамина и превратить урацил в цитозин, а дУДФ в дЦДФ, а дЦДФ в дЦТФ. дУДФ можно превратить в дТДФ (тимидиндифосфат), а потом в дТТФ. УДФ можно превратить в УТФ, а тот с аммиаком от глутамина в ЦТФ. В общем подробности на рисунке ниже.
Чуть подробнее должен сказать, как рибонуклеотиды превращаются в дезоксирибонуклеотиды. Фермент рибонуклеотидредуктаза вырывает кислород из 2-го положения рибозы, цепляет на него два водорода от сульфгидрильных групп тиоредоксина, получается дезоксирибонуклеотид и вода, однако в тиоредоксине образуется дисульфидная связь, но эту проблему с помощью НАДФН2 решает тиоредоксин-редуктаза. Всё на рисунке. Система работает и для пуриновых, и для пиримидиновых нуклеотидов.
123

Пуриновые нуклеотиды
Теперь обсудим пуриновые нуклеотиды.
Основой пуриновых оснований является пурин, основные основания, которыми мы оперируем – аденин и гуанин, но в катаболизме этих оснований мы встретимся с гипоксантином и ксантином, так что их бы тоже надо знать. Источники атомов в пурине указаны на рисунке, реакции его синтеза учить не надо – их 10 и они очень сложные – в программу их не включают, а вот синтез уже самих нуклеотидов знать нужно. Сначала берётся ФРПФ, от него отщепляют пирофосфат и начинают строить на этом месте азотистое основание, получается инозинмонофосфат ИМФ (придумали такое название, потому что буква «Г» для аббревиатур уже занята гуанином). С помощью аспаргиновой кислоты и ГТФ из ИМФ получают АМФ (в гипоксантине кислород меняют на аминогруппу, получая аденин), также можно с помощью АТФ и глутамина приделать аминогруппу (приделать, а не заменить) к гипоксантину и превратить его в гуанин. А дальше можете прибавлять им ортофосфаты и/или превращать в дезоксирибонуклеотиды.
Уловите разницу – пиримидины синтезируют, а потом надевают на
рибозо-5-фосфат от ФРПФ, а пуриновые строят сразу на рибозо-5- фосфате.
Ферменты синтеза нуклеотидов могут являться мишенями для действия противовирусных и противоопухолевых препаратов – если заблокировать эти ферменты, то не будут синтезироваться нуклеотиды и вирусам с опухолевыми клетками будет не из чего строить новые ДНК для размножения, да и РНК тоже. Примеры. 5-фтор-урацил ингибирует
124

тимидилатсинтетазу – нет тимина, а значит, не из чего строить ДНК. Метатрексат – структурный аналог фолиевой кислоты, ингибирует дигидрофолатредуктазу, что не позволяет синтезировать пуриновые нуклеотиды.
Катаболизм пуриновых нуклеотидов Разберём
катаболизм пуриновых нуклеотидов.
125
Начнём с АМФ, первым делом нуклеотидаза убирает гидролитически ортофосфат, затем адениловая дезаминаза с помощью воды меняет аминогруппу на кислород. Фосфора и аммиака нет – можно разрушать нуклеозид, что и делает нуклеозидфосфорилаза – она действует, как гидролаза, только вместо воды ортофосфат – образуется рибозо-1-фосфат и гипоксантин. На гипоксантин действует ксантиноксидаза – она складывает кислород и воду, чтобы образовать перекись и атом кислорода, который она презентует гипоксантину, делая его ксантином, потом повторяет манипуляцию, только в другом участке молекулы, образуя, в итоге, мочевую кислоту – конечный продукт обмена пуриновых нуклеотидов. Заметьте, что этот фермент использует ФАД, железо и кобальт (на дороге не валяется). У ГМФ похожая тактика – лишается ортофосфата нуклеотидазой, лишается аминогруппы с помощью гуанилатдезаминазы через воду. Аммиака и фосфора нет – запускаем нуклеозидфосфорилазу – получаем ксантин, на него действует ксантиноксидаза, получаем мочевую кислоту. Т.е. в любом случае мочевая кислота, которая очень плохо растворяется в воде, если её будет образовываться слишком много (много пуринов поступает в катаболизм), её концентрация превысит предел растворимости и мочевая кислота станет оседать в суставах в форме кристаллов с невероятно острыми концами. Макрофаги захотят растворить этот кристалл, но наткнутся на его острые края, разорвутся и выпустят все гидролитические ферменты в окружающие ткани, ферменты будут разрушать всё на своём пути – начнётся асептическое (безбактериальное) воспаление суставов – тофусы – нереально болезненная вещь. Помимо этого, мочевая кислота образует камни в почках (ураты). Всё это называется подагрой. Существует система, которая не даёт перегружать путь катаболизма пуринов, дабы мочевой кислоты не вырабатывалось слишком много – реутилизация пуринов.
126

По сути азотистое основание надевается на рибозо-5-фосфат от ФРПФ, получается безобидный нуклеотид, азотистое основание избегает катаболизма. Аденинфосфорибозилтрансфераза работает только с аденином, если она сломается, то вам грозит уратная почечно-каменная болезнь. Гипоксантин-гуанин-фосфорибозилтрансфераза работает с гипоксантином и гуанином, если она ломается у взрослых, наступает подагра (камни в почках + тофусы), а у детей болезнь Лёши-Нихона (симптомы те же, но с рождения и тяжелее, живут до 10-ти лет). Подагра чаще встречается у мужчин, особенно если они едят много икры, рыбы и мяса (много пуринов там), а вообще это заболевание генетически обусловлено (мультифактериальное). Патогенез и симптоматику я описал, каково лечение? Во-первых, диета, исключающая продукты, богатые пуринами, вовторых, приём аллопуринола, который обратимо конкурентно ингибирует ксантиноксидазу, чтобы гипоксантин и ксантин не превращались в мочевую кислоту, а просто выводились с мочой (они более растворимы, чем мочевая кислота). Болезнь Лёши-Нихона лечится так же.
Буковкой «Р» я обозначаю ортофосфат, когда не хочу писать его в развёрнутом виде. В жкт нуклеозиды разрушает водой нуклеозидаза, а в клетках нуклеозиды разрушает нуклеозидфосфорилазы ортофосфатом.
127

Катаболизм пиримидиновых нуклеотидов
И последнее в теме нуклеинового обмена – катаболизм пиримидинов, он, в отличии от пуринов, идёт до конца, т.е. до воды, углекислого газа и аммиака.
Логика здесь похожа на пуриновый катаболизм в начале – долой амины и фосфор. Вот и здесь ЦМФ лишается ортофосфата под действием нуклеотидазы, потом цитозиддезаминаза меняет его аминогруппу на кислород, получается урацидин, который также получается, когда на УМФ действует нуклеотидаза. Дальше на урацидин действует пиримидиновая нуклеозидфосфорилаза, оставляя рибозо-1-фосфат и делая урацил. На урацил действует дигидроурацилдегидрогеназа, которая устраняет двойную связь, присоединяя водороды от НАДФН2, получается дигидроурацил (т.е. с двумя «лишними» водородами), далее в результате двух реакций он теряет аммиак и углекислый газ, остаётся 3-аминопропилат (промежуточный продукт), который способен распадаться до аммиака, углекислого газа и воды. Итак, ЦМФ и УМФ разложили до конечных продуктов.
128

Катаболизм дТМФ
Теперь дТМФ. Его катаболизм.
По сути всё так же, как и у УМФ, вот только обязательно помните, что при тимине у нас не рибоза, а дезоксирибоза! Второе важное отличие – промежуточным продуктом является не 3-аминопропилат, а 3-амино-2- метил-пропилат, но распадается он на те же продукты. Название продуктов и ферментов соответствующие – см. рисунок выше.
Поздравляю, тема нуклеинового обмена окончена!
129
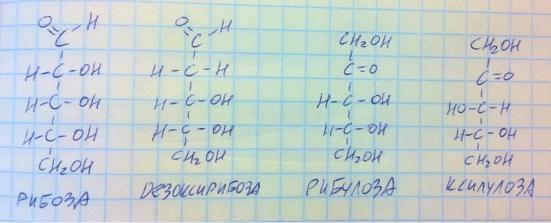
Углеводный обмен
Прежде всего, надо определиться с тем, кто такие углеводы. Углеводы
– это органические вещества, соответствующие общей формуле Cn(H2O)m, хотя есть небольшие исключения вроде дезоксирибозы, также углеводы являются многоатомными спиртами и карбонильными (кетоны/альдегиды) соединениями одновременно. Могут полимеризоваться. Вот это по сути полное представление о том, что такое углеводы. Теперь к классификации. Первый признак классификации – количество атомов углерода в мономере, их бывает от 3-х до 10-ти, называют их, соответственно триоза, тетроза, пентоза, гексоза, гептоза и так далее, но нас больше всего интересуют пентозы (5) и гексозы (6). Второй признак классификации – карбонильная группа – она бывает кетонной и альдегидной, следовательно, углеводы разделяются на кетозы и альдозы. Третий признак классификации полимерность – выделяют моносахариды, олигосахариды (2-10 мономеров, отдельно рассматривается подкласс дисахаридов), полисахариды (от 10 и больше, обычно существенно больше, они в свою очередь подразделяются на гомополимеры и гетерополимеры). И имейте ввиду, что в нашем организме все углеводы в D-формах. Поговорим конкретнее о представителях, которых нам надо знать.
Моносахариды. Пентозы. Необходимо знать 4 представителя: рибозу, дезоксирибозу, рибулозу, ксилулозу.
Как видите, рибоза и дезоксирибоза являются альдозами, а рибулоза и ксилулоза – кетозами. Как запомнить эти формулы? Они большие и их несколько. Есть простой вариант – запомнить одну формулу и отличия от неё других формул. Запомните рибозу – сверху альдегид, снизу CH2OH (это у всех моносахаридов), а все гидроксильные группы смотрят направо, углеродов 5. Дезоксирибоза отлична от рибозы тем, что у второго углерода нет гидроксильной группы – только водород. Рибулоза от рибозы отличается тем, что 1-й и 2-й углероды поменялись карбонильной и гидроксильной
130