

4.2. Метаболизм липидов
Под действием озона происходит снижение содержания липидов, которое может происходить за счет ингибирования их биосинтеза. В результате наблюдается уменьшение содержания всех жирных кислот, причем в наибольшей степени снижается количество пальметиновой и линоленовой кислот. Это снижение содержания жирных кислот и изменение их состава связано с нарушением жизнеспособности клеток.
Озон нарушает способность метаболизировать (I14C) ацетат или ацетил-Ко-А (ацетилкоэнзим А) в биосинтезе липидов. В частности он ингибирует синтез гликолипидов в большей мере, чем фосфолипидов, и способствует образованию глютатиона. Растет и число стерольных гликозидов, но снижается количество свободных стеролов. При низких концентрациях озона (> 0,5 часть/млн) и при продолжительном действии происходит увеличение свободных стеролов и уменьшается число стероидных эфиров и гликозидов.
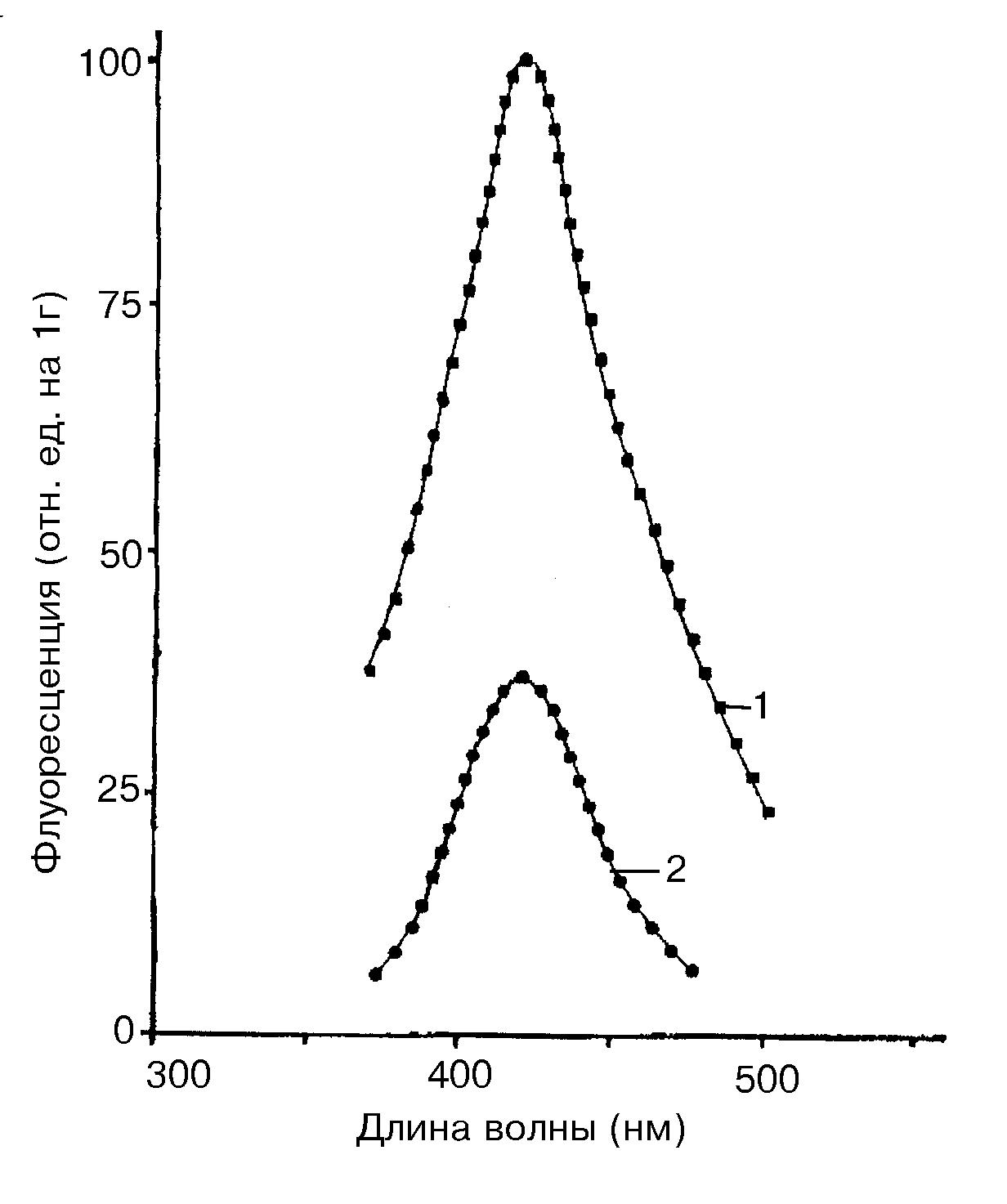
Липиды в клетках защищены от окисления белковым слоем или наличием большого количества антиоксидантов, но эта защита не может быть полной. Поэтому в клетках часто имеет место перекисное окисление, вызванное пероксидами и свободными радикалами, образующимися при атаке озона. В результате образуются малондиальдегид, флуоресцирующие Шиффовы основания и даже пигмент старения липофусцин. Липофусциновые пигменты впервые были найдены у животных при экстремальных воздействиях. Они накапливаются, и появляется характерная их флуоресценция в видимой части спектра. Появление липорастворимых флуоресцирующих связано с взаимодействием продуктов перекисного окисления липидов (в частности малондиальдегида) с соединениями, содержащими, свободные аминокислоты. Формирование желтого пигмента старения - липофусцина связывают с превращением каротиноидов. Подобные продукты образуются и в растениях при действии озона, например в липидных фракциях семян сои Glycine soja L. и кукурузы Zea mays L. , флуоресцирующие в области 420-440 также в нм (Рис 19).
|
Рис.19. Спектры флуоресценции липофусциновых фракций, образующихся из семян сои Glycine soja L. экспонированных на воздухе, содержащем 1,5 часть/млн. озона в течение 100 часов при комнатной температуре (1) и на воздухе без озона (2) [Brooks, Csallany, 1978].
|
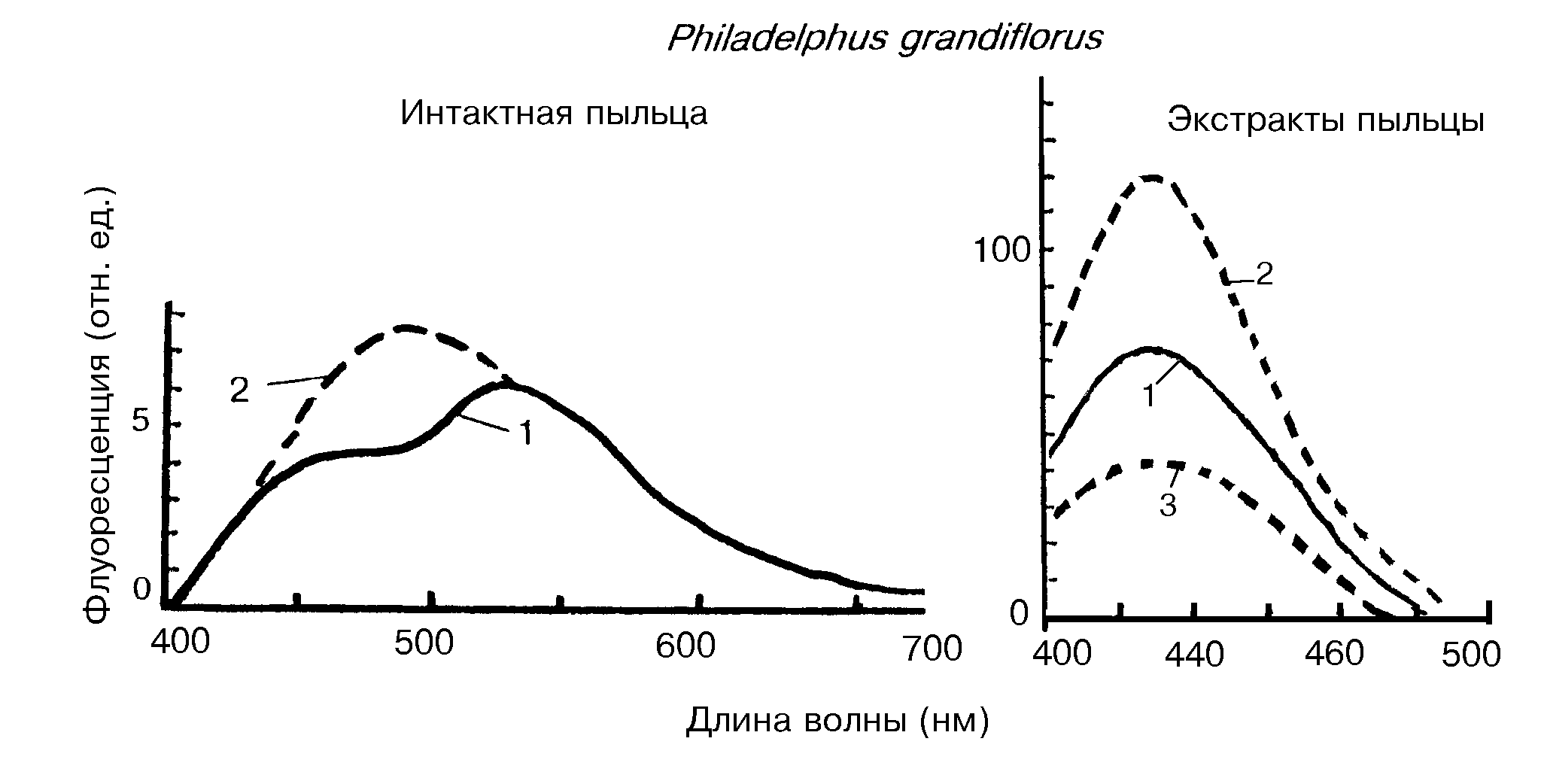
Озон стимулировал синтез липофусцина в 2,7 раза. Липофусцин обнаружен и в созревающих плодах и стареющих листьях. Образование липофусцино-подобных пигментов под влиянием озона обнаружено также в пыльце, обогащенной каротиноидами (рис.20).
Анализ хлороформ/этанольных экстрактов из пыльцы обнаружил пигмент с максимумом 430 нм, который сходен с липофусцинами. У объектов, где мало каротиноидов, и они экранировались другими пигментами, или пигменты отсутствовали как у подорожника Plantago major, липофусцин не образовывался.
4.3.Углеводный метаболизм. Дыхание и фотосинтез
Биохимические изменения под действием озона часто выражаются в сдвигах углеводного метаболизма, что заметно по содержанию растворимых сахаров и других углеводов, прежде всего у растений. Оно может, как увеличиваться, так и уменьшаться. Например, в листьях сои Glycine soja L. и хвое сосны Pinus L. после
|
Рис.20. Образование новых флуоресцирующих продуктов под действием озона у пыльцы Philadelphus grandiflorus [Roshchina, Karnaukhov,1999].Слева - спектры флуоресценции интактной пыльцы в контроле (1) и после хронического озонирования (2) в течение 100 часов по 3 часа в день (Общая доза озона 5 часть/млн.). Справа-спектры флуоресценции хлороформ/этанольных экстрактов (1:1 по объему) из этой же пыльцы , необработанной (1) и после обработки озоном без очистки (2) или очистки флуоресцируюшего компонента Rf 0,2 с помощью тонкослойной хроматографии (3) |
экспозиции в озоне (0,49 часть/млн.) немедленно уменьшается количество растворимых сахаров. При этом содержание крахмала не изменяется, что свидетельствует об уменьшении пула растворимых сахаров в результате угнетения фотосинтеза. При однократном воздействии озона вслед за уменьшением следует возрастание содержания восстановленных сахаров. Это увеличение может быть результатом нарушения транслокации сахаров или ингибирования синтеза крахмала. Под действием озона происходит и снижение включения радиоактивной метки в простые растворимые сахара и ускорение включения в сахарофосфаты в связи с усилением дыхания. Прежде всего, у растений это определяется соотношением скоростей дыхания и фотосинтеза.
Фотосинтез является более чувствительной реакций к озону, чем дыхание для растений. Скорость фиксации углекислого газа начинает быстро снижаться после 60 мин. экспозиции в озоне и в конце воздействия (через 120 мин.) ингибируется на 75%. Считается, что озон нарушает функции тилакоидных мембран, поскольку ингибирует электронный транспорт в обеих фотосистемах (ФС I и ФС II) и сопряженный синтез АТФ. Увеличение концентрации озона первоначально повреждает донорную сторону ФС II (система разрушения воды), а лишь затем ингибирует электронный транспорт от ФС I к ФС I. В целом этот газ повреждает структуру хлорофилл-белкового комплекса. При использовании высоких концентраций озона хлорофилл разрушается очень быстро, при этом хлорофилл "б" разрушается быстрее хлорофилла "а". Снижение фотосинтеза может быть обусловлено и непосредственным действим на карбоксилазы, участвующие в фотосинтетической фиксации СО2. Индуцированная озоном грануляция стромы хлоропластов происходит за счет окисления SH-групп рибулезодифосфат карбоксилазы (РДФ-карбоксилазы). Ингибирование этого фермента происходит, например у овса Oryza sativa, вскоре после 2 часовой экспозиции в озоне (0,12. мкл/л).. Полагают, что решающим фактором снижения поглощения СО2 является уменьшение эффективности карбоксилирования триозофосфатов,
Озон может в целом ингибировать фотосинтез, стимулировать, и ингибировать дыхание организмов. Эффект зависит от объекта исследования, концентрации и продолжительности действия озона. В листьях растений часто наблюдается повышение интенсивности дыхания при одновременном снижении фотосинтеза. В целом, дыхание является процессом, менее чувствительным к озону по сравнению с фотосинтезом.
Изменения дальнейшего метаболизма углеводов происходят за счет действия О3 на энзиматическую активность на отдельных этапах образования сахаров. На рис.21 показаны участки действия озона, связанные с модификацией соответствующих ферментов. Под действием озона изменяется активность АТФ-азы и ферментов превращения фосфорилированных сахаров. Озон подавляет ферментные системы синтеза целлюлозы и крахмала, ингибирует гликолитический путь обмена и стимулирует пентозофосфатный путь. При низких концентрациях озона (0,05 мкл/л или часть/млн.) ингибируется гидролиз крахмала за 2-6 часов. Наблюдаемый эффект, по-видимому, происходит за счет ингибирования фосфорилазы. У растений в пользу подавления ферментной системы синтеза
|
Рис.21. Мишени для озона в углеродном метаболизме |
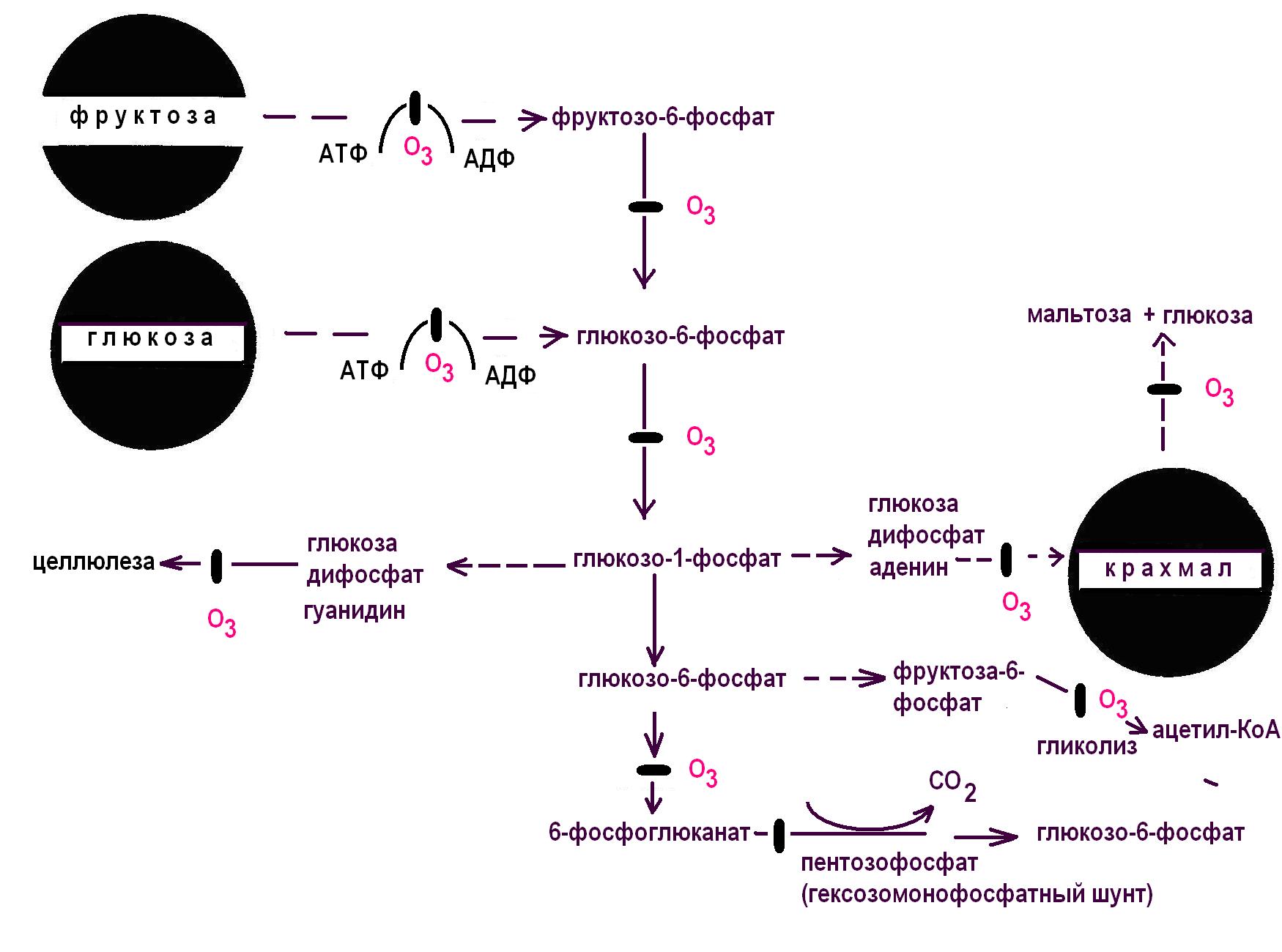
целлюлозы свидетельствуют опыты, в которых обнаружено, что при концентрации озона 116 часть/млн. уменьшается содержание целлобиозы в 2 раза и снижается содержание полисахаридов. Предполагается, что озон подавляет синтетазы (фосфоглюкомутазу), необходимые для синтеза целлюлозы. Напротив, целлюлазная активность увеличивается, как пероксидазы и менее значительно лактатдегидрогеназы. Быстрое увеличение целлюлазной и амилазной активности под действием озона указывает на быстрое разрушение клеточной стенки и запасного крахмала. Активность целлюлазы при этом может увеличиваться до 400% по сравнению с контролем.