

5.2. Растения как биоиндикаторы озона
Поскольку видимые повреждения лучше всего заметны на растениях, то основные определения повреждающих доз озона разработаны именно на растительных организмах (см. Приложение 1). Биологический эффект определенной дозы озона чаще всего определяется по повреждению листьев растений при хроническом или остром воздействии этого газа. Взаимосвязь между концентрацией и временем действия озона не определяется точно одной пороговой дозой, поскольку часто одна и та же степень повреждения листа, например, может быть вызвана острой дозой 0,1 часть/млн. (196 мкг/м 3) и выше всего за один час или хронической небольшой концентрацией газа 0,05 частей на млн (98 мкг/м 3) за 10 часов. Очень важен вопрос о пороговой дозе, вызывающей видимые повреждения у чувствительных видов. У чувствительного к озону сорта табака Bel-W3 листья повреждаются при пороговой дозе 0,05 часть/млн за 4 часа (210-9 моль/л за 4 часа). Хотя этот порог может варьировать в зависимости от условий внешней среды, он принят за основную пороговую величину. Построена шкала развития повреждений в % поврежденной листовой поверхности в зависимости от времени фумигации. Линейная кривая доза-эффект для озона возможна только в радиусе 1/4 мили вокруг воспринимающего озон растения в течение месяца. В основном, эти кривые носят нелинейный характер.
Порог чувствительности зависит и от индивидуальных особенностей растительного организма. В целом очень чувствительные растения повреждаются при концентрациях озона 0,05-0,1 часть/млн. Нормальный же уровень содержания О3 в воздухе согласно международным стандартам должен быть 0,08 частей на млн (157 мкг/м3) за 1 час не более раза в году. В урбанизированных районах он гораздо выше.
Чувствительность видов и даже сортов одного и того же вида растений к озону, определяемая по повреждению вегетативных клеток, очень различна. Как правило, подавляющее большинство растений толерантно к озону и повреждения проявляются только в случае сильного атмосферного загрязнения. Однако незначительное число видов растений очень чувствительно к О3 и повреждаются при концентрациях озона лишь слегка превосходящих фоновые. Именно эти растения наиболее ценны для биологической индикации. Список индикаторных видов растений приведен в приложении 2. Кроме того, чувствительность растений к озону зависит от многих внешних факторов: эдафических, климатических, атмосферных и биотических , поэтому при биоиндикации повреждений надо их учитывать (Приложение3).
5.2.1.Ранняя диагностика озоновых повреждений по биохимическим и физиологическим критериям
Повреждение клеток озоном, как и многими другими факторами, проявляется в первую очередь на биохимическом и физиологическом уровнях. Это изменение в активности ферментов, содержании пигментов, увеличении образования этилена, снижении биосинтеза АТФ, снижении скорости фотосинтеза, нарушении водообмена. Подобные реакции часто наблюдают раньше видимых озоновых повреждений листьев. Поэтому предлагается использовать эти более быстрые реакции как индикаторы озоновых эффектов (рис. 24).
Раньше того, как повреждение проявится во внешних признаках - хлорозах, некрозах и т.д., можно обнаружить подобные физиологические и биохимические изменения растений.
|
Рис. 24. Схема возникновения и распространения озоновых нарушений биохимических и физиологических процессов в зависимости от дозы озона |

Биохимические реакции растений на озон имеют разные пороги чувствительности in vivo. При самых низких концентрациях озона 0,05 часть/млн уже наблюдается ингибирование гидролиза крахмала. Затем по мере повышения концентрации этого газа до 0,2-1,0 часть/млн происходит ингибирование фотосинтеза, понижение концентрации хлорофилла b, снижение уровня НАДН и фосфорилирования, ингибирование дыхания или, наоборот, его стимуляция, а также увеличение содержания свободных аминокислот. Образование малондиальдегида определяется уже после развития симптомов повреждения листа уже при фумигации 0,25 часть/млн озона в течение 3х часов, например у фасоли Phaseolus vulgaris, хотя у других видов это заметно при более высоких концентрациях 0,8-1,0 часть/млн в течение 5 часов.
При продолжительных воздействиях озона на клетки происходят изменения основного метаболизма. В азотном обмене – это усиленный биосинтез определенных белков-ферментов (глюканаз, хитиназ, пероксидаз) и повышение активности антиоксидантных ферментов супероксиддисмутазы, пероксидазы, каталазы, глютатионредуктазы. Для углеводного обмена характерно повышение концентрации растворимых сахаров, а для липидного обмена – изменение состава липидов и перекисное их окисление с образованием малонового диальдегида, а позднее – пигмента старения липофусцина. Малоновый диальдегид, связываясь с аминокислотными группами белков, образует Шиффовы основания, которые легко идентифицировать по характерной их флуоресценции в области 440-560 нм. Особенно интересны изменения вторичного обмена, поскольку образуются защитные соединения, как общие для всех организмов, такие как полиамины, некоторые фенолы, а для растений еще и фенольные пигменты – антоцианы, стилбены и др., а также - углеводороды, например стрессовый этилен.
Высокие дозы озона вызывают глубокие изменения клеток, которые можно диагностировать по увеличению проницаемости мембран и вымыванию ионов, скорости общего фотосинтеза и выцветанию хлорофилла, а также по изменению метаболических пулов углеводов, липидов, терпенов,фенолов и полиаминов.
Для ранней диагностики изменений клеток, вызванных озоном, могут использоваться как общие реакции растительного организма (образование стрессового этилена, изменение активности и содержания ряда ферментов, флуоресценция хлорофилла, изменение состава пигментов и др.), так и реакции специализированных клеток (автофлуоресценция секреторных клеток, прорастание пыльцы и спор и т.д.).
Наиболее общие реакции на озон связаны с выделением метаболитов, изменением их состава, а также активности ферментов.
Стимуляция выделения этилена. При стрессах биосинтез этилена увеличивается в несколько раз раньше нарушения проницаемости мембран и вымывания каких-либо соединений, поэтому этот показатель тоже может использоваться для диагностики.
Изменение активности и биосинтеза ферментов. Ферменты, как многие белки, быстро взаимодействуют с озоном, и в результате этого изменяется их активность, а длительное воздействие озоном приводит к нарушению на генетическом уровне - изменяется и биосинтез ферментов (репрессируется или активируется). Чаще всего наиболее заметно активируется функция оксидаз, таких как пероксидаза. Именно нарастание активности пероксидазы и (или) усиленный биосинтез ее различных изоформ при озоновом стрессе могут быть наиболее четким показателем ответа растения на озон. Отмечено, что изменение активности пероксидаз происходит раньше появления внешних признаков повреждения. Кроме того, многие реакции, катализируемые пероксидазой, сопровождаются образованием интенсивно окрашенных соединений. Это существенно облегчает обнаружение пероксидазной активности. Часто под влиянием озона подавляется образование ферментов синтеза целлюлозы и крахмала, но увеличивается синтез и активность целлюлазы и амилазы, ферментов распада клеточной стенки и крахмала. Активность целлюлазы может увеличиваться до 400%. Таким образом, активности как пероксидаз, так и и целлюлаз могут быть тестами на озоновые повреждения.
Увеличение проницаемости мембран и вымывание ионов. Озон значительно увеличивает проницаемость мембран для ионов К+, Са2+, Мg2+, и их концентрация в экстраклеточной среде увеличивается, что является показателем заметного повреждения мембранных структур.
Скорость фотосинтеза и изменения состава пигментов. Фотосинтез может использоваться как тест-процесс для ранней диагностики озонового стресса уже при концентрации 310-7М О3. Чаще всего под влиянием озона изменяются скорость фотоассимиляции, уровень флюоресценции хлорофилла и соотношение форм хлорофилла a/b (с усилением озонового стресса обычно снижается это отношение). Самый быстрый ответ на изменение состояния хлорофилла в листе - это его флуоресценция в красной области спектра. Озон стимулирует флуоресценцию хлорофилла. Индукция флуоресценции хлорофилла (так называемый эффект Каутского) вызванная озоновым (0,3-0,5 мкл/л) повреждением листьев фасоли Phaseolus vulgaris наблюдается по крайней мере за 20 часов до видимого некроза листьев. Первый видимый эффект изменения флуоресценции связан с повреждением ферментов, участвующих в разложении воды, после чего следует ингибирование электронного транспорта между фотосистемами.
Изменение метаболических пулов. Накопление одних метаболитов и снижение уровня других под действием озона могут быть замечены раньше видимых изменений в растениях. Чаще всего индикаторами озонового стресса могут быть пулы растворимых сахаров, поскольку этот газ подавляет ферментные системы синтеза целлюлозы и крахмала, в результате чего в клетках снижается содержание полисахаридов. Пул растворимых сахаров или аминокислот снижается, из-за снижения фотоассимиляции и роста интенсивности дыхания. Соотношение содержания насыщенных и ненасыщенных жирных кислот, также может быть показателем озонового токсикоза. Снижение содержания липидов обычно идет за счет ингибирования их синтеза. Особенно четкими в качестве индикаторов озонового стресса являются признаки перекисного окисления липидов. В этом случае количество ненасыщенных жирных кислот уменьшается, а содержание малонового альдегида возрастает.
Наиболее специфичными признаками для определения именно озонового повреждения являются изменения пулов вторичных метаболитов. Чаще всего показателем при загрязнении оксидантами будет изменение концентрации монотерпенов в хвое у голосеменных растений, например, у Pinus abies при ее пожелтении. Накопление или исчезновение некоторых фенолов может быть достоверным показателем загрязнения среды именно оксидантами. Показателем озонового стресса является и накопление полиаминов - путресцина, спермина, спермидина.
Реакции на озон специализированных клеток, таких как секреторные клетки (железы вегетативных тканей, генеративные клетки – пыльца, вегетативные микроспоры), могут рассматриваться как более специфичные по сравнению с выше изложенными реакциями, отмеченными и для многих других видов стресса. К подобным процессам относят автофлуоресценцию секреторных клеток и скорость прорастания пыльцы и вегетативных микроспор.
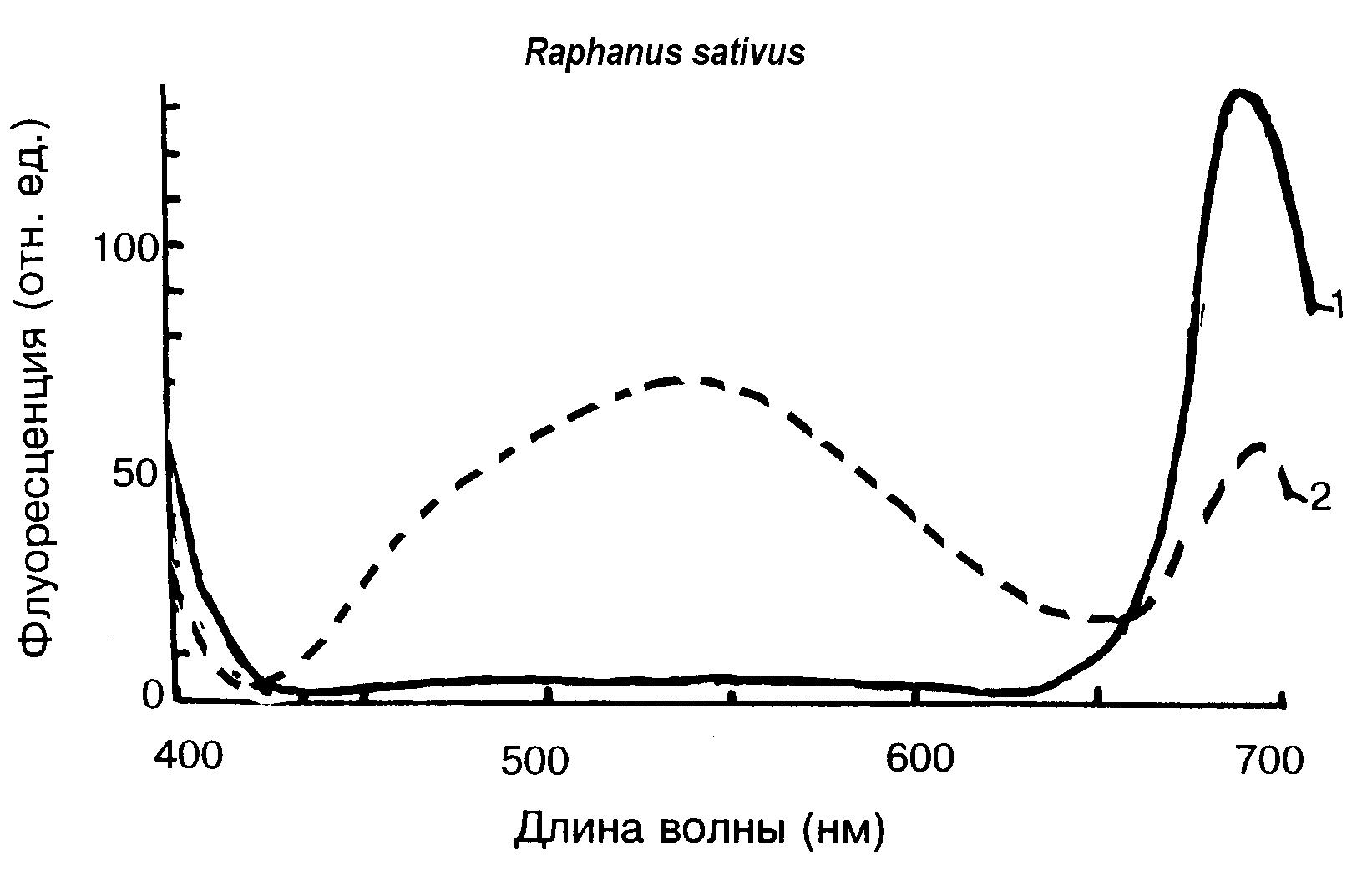
Автофлуоресценция секреторных клеток. Автофлуоресценция секреторных клеток растений может служить показателем ранней диагностики озоновых эффектов. Метод флуоресценции является простым, быстрым, не вызывающим деструкции ткани. Особенно полезным является микроспектрофлуориметрия, регистрирующая спектры флуоресценции с индивидуальных клеток. Например, после фумигации озоном проростков редиса (Рис.25) в их листовых секреторных клетках (железистых секреторных волосках) ранее слабо флуоресцирующих, появляется отчетливое желтое свечение. На ассимилирующей поверхности помимо изменений флуоресценции хлорофилла под влиянием озона отмечаются сдвиги максимумов в голубой и оранжевой области спектра и появление голубых флуоресцирующих пятен на поверхности листьев. Они могут быть связаны как с изменением состояния фенольных и терпеноидных соединений, имеющих двойные связи, так и с новообразованием пигмента старения липофусцина.
|
Рис.25. Появление флуоресцирующих соединений в спектрах флуоресценции секреторных волосков листьев проростков редиса. 1- без обработки озоном, 2 – после фумигации озоном в общей дозе 0,15 мкл/л за 3 часа |
В последние годы также как чувствительный тест на озон рассматриваются изменения автофлуоресценции пыльцы семенных растений и вегетативных микроспор несеменных растений. Показано, что пигменты на поверхности пыльцы или вегетативной микроспоры определяют ее устойчивость к О3. По сдвигам максимумов в спектрах флуоресценции пыльцы, можно судить о степени ее повреждения.
Механизм действия озона на пыльцу рассмотрен на основании данных о взаимодействии озона и пероксидов с поверхностными компонентами спорополленина и эти процессы смоделированы и в искусственных системах. В качестве тест-реакции измеряли автофлуоресценцию пыльцы, которая обусловлена составом компонентов спорополленина. Фумигация озоном резко изменяет интенсивность свечения и положение максимумов в спектрах флуоресценции (Рис.26). У пыльцы пассифлоры Passiflora coerulea или красоднева Hemerocallis fulva максимум флуоресценции после обработки озоном сдвигался в коротковолновую область, интенсивность же свечения резко возрастала. Другие виды демонстрировали или снижение интенсивности свечения, как у сосны Pinus sylvestris, или ее увеличение и появление новых максимумов, как у ежи сборной Dactylis glomerata или изменяется как
|
Рис.26. Спектры флуоресценции пыльцы различных растений до (1) и после фумигации озоном 0.5 частей на млн. в течение 3 часов (2) |
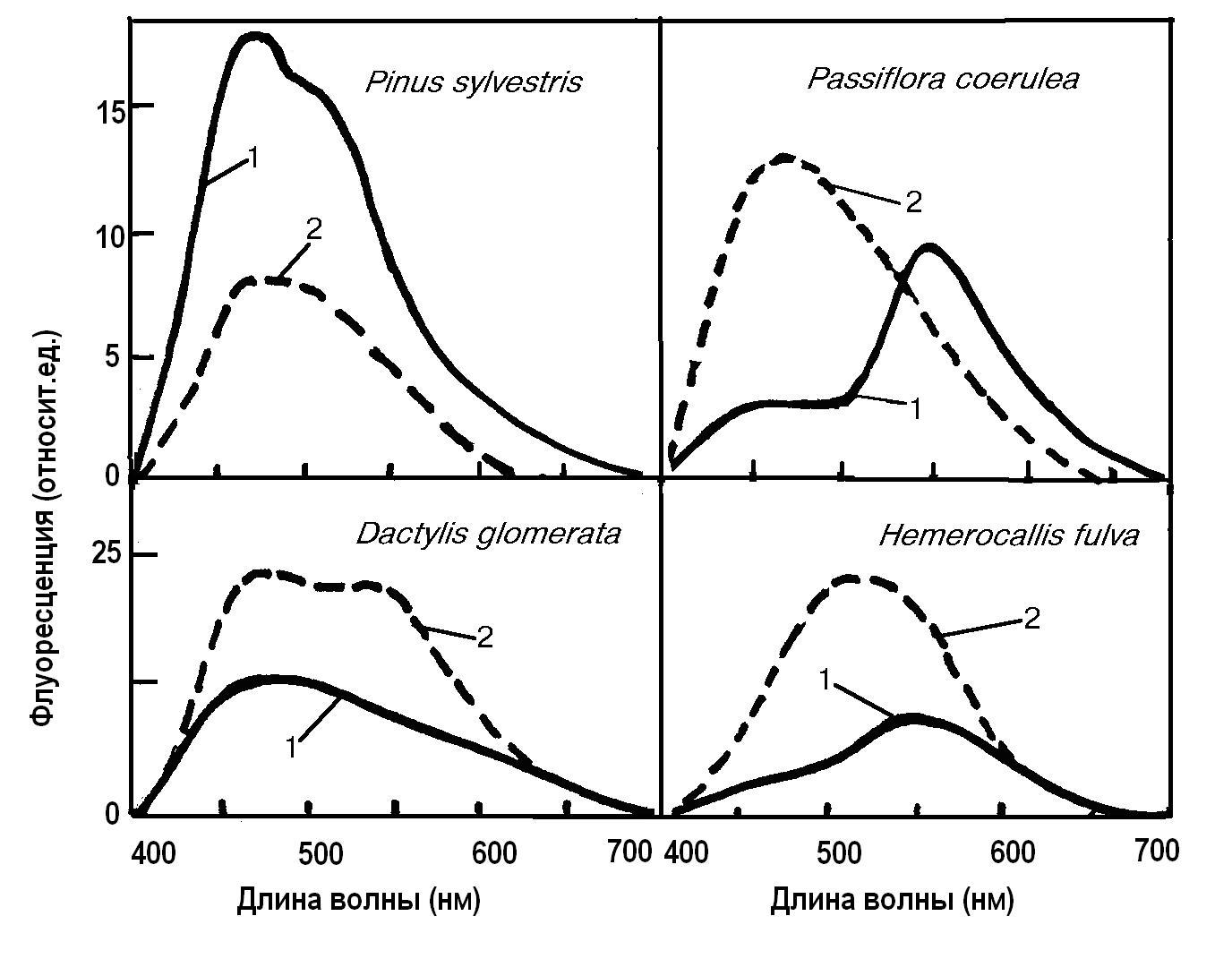
интенсивность свечения, так и положение максимумов в спектре флуоресценции пыльцы Passiflora coerulea. У последнего вида максимум флуоресценции после обработки озоном сдвигался в коротковолновую область, интенсивность же свечения резко возрастала. Таким образом, озоновые повреждения пыльцы выражаются в изменении спектра и интенсивности флуоресценции. Оба эти показателя зависят от вида растения и концентрации озона. Возможно, что отмеченные изменения являются наиболее ранними показателями повреждения, поскольку отмечаются всего после нескольких часов фумигации озоном.
Цвет и интенсивность автофлуоресценции вегететивных микроспор споровых растений также резко изменяются в присутствии небольших ( ~ 0.05 мкл/Л) концентраций озона. Например, появляется желтое свечение клеток, раньше, чем отмечают признаки повреждения клеток.
Прорастание пыльцы и рост пыльцевой трубки. Озон оказывает различное влияние на прорастание пыльцы in vitro и in vivo. В зависимости от концентрации озона наблюдается снижение этого процесса и угнетение роста пыльцевой трубки. Например, рост пыльцевой трубки на агаровых дисках с 10% сахарозой полностью блокируется, если развитие пыльцы происходит после 5-ти часовой фумигации озоном (1 часть/млн.). Более низкие концентрации озона вызывали 40-50% снижение прорастания пыльцы и на 50% элонгацию пыльцевой трубки у чувствительного к озону сорта табака Bel W3.
Существует связь чувствительности пыльцы к озону с пигментацией пыльцевых зерен. Это показано в опытах с прорастанием пыльцы с разным составом экзины, поверхностного слоя оболочки: чубушника Philadelphus grandiflorus (преобладают азулены и каротиноиды), гиппеаструма Hippeastrum hybridum, пыльцы, не содержащей таких пигментов, и бесцветной пыльцы подорожника Plantago major. При увеличении времени фумигации у всех трех образцов пыльцы индекс прорастания в % к контролю снижался (рис.27). Однако у пыльцы чубушника Philadelphus grandiflorus, обогащенной каротиноидами (антиоксидантами), способность к прорастанию сохранялась гораздо дольше, чем у пыльцы других видов, не содержащей таких пигментов, или бесцветной. Пыльца, лишенная или содержащая мало пигментов, наиболее чувствительна к
|
Рис. 27. Влияние озона на индекс прорастания пыльцы разных видов растений в % от контроля [Рощина, Мельникова, 2001]. Бар -средняя ошибка , при n=4. 1 - Philadelphus grandiflorus ; 2 - Hippeastrum hybridum; 3- Plantago major |
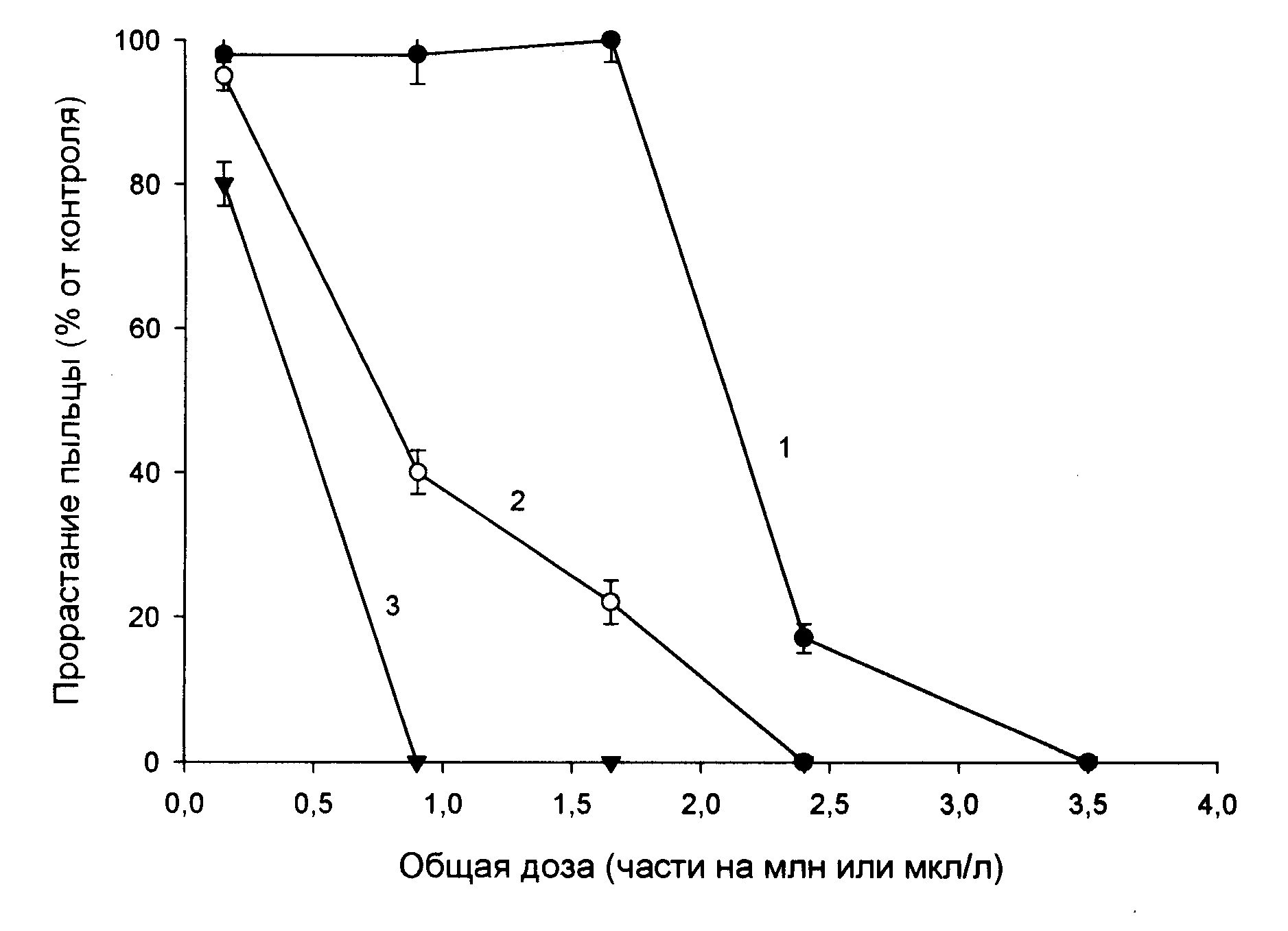
озону и может использоваться для его индикации.
Прорастание вегетативных микроспор споровых растений. Вегетативные микроспоры растений, размножающихся спорами, могут быстро прорастать в присутствии низких концентраций озона, но высокие его концентрации ингибируют процесс