

Вирусы, содержащие двухцепочечную рнк
Двухцепочечный сегментированный (10 сегментов) геном реовируса транскрибируется вирионной полимеразой внутри частично «раздетого» капсида, через открытые вершины которого выходят 10 мРНК — плюс-транскрипты 10 генов Молекулы мРНК выполняют две функции. Во-первых они транслируются, обеспечивая синтез вирусных белков, .и, во-вторых, они включаются (по одной молекуле мРНК каждого из 10 генов) в состав частиц-предшественниц. Каждая мРНК служит в этих частицах матрицей для синтеза комплементарной цепи, что ведет к образованию двухцепочечных сегментов генома.

Размножение реовирусов.
Геномы днк-содержащих вирусов
Геномы ДНК-содержащих вирусов можно разделить на четыре группы. Геномы папова-, адено- и герпесвирусов транскрибируются и реплицируются в ядре, поэтому они могут использовать для синтеза мРНК ферменты транскрипции хозяина. Этим объясняется инфекционность ДНК указанных вирусов.
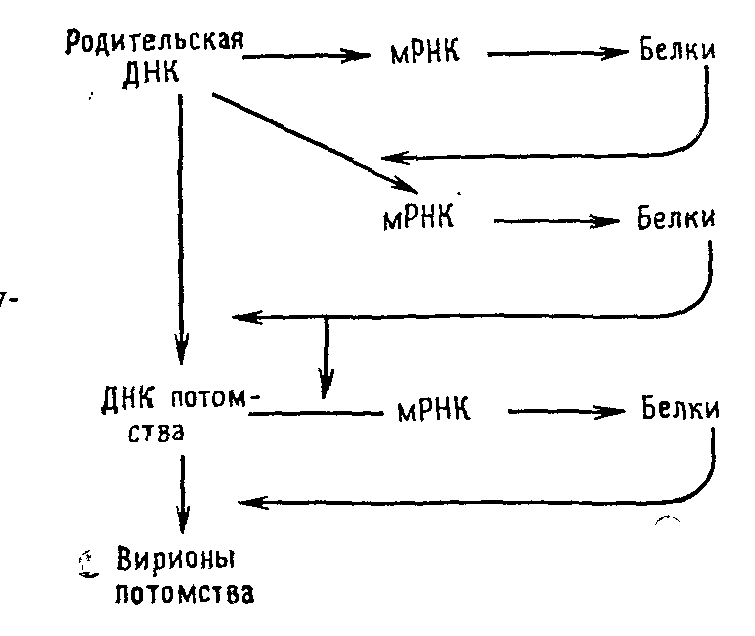
Размножение герпесвирусов (вируса простого герпеса).
У паповавирусов транскрипционная программа состоит по крайней мере из двух циклов, а у герпес- (рис. 5.7) и аденовирусов по крайней мере из трех. В каждом случае структурные или вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции.
ДНК-содержащие вирусы, репродукция которых происходит в цитоплазме (поксвирусы и иридовирусы) составляют вторую группу. Хотя ДНК поксвирусов удается обнаружить в ядре, по крайней мере начальные события транскрипции и большая часть остальных событий цикла репродукции происходят в цитоплазме. Начальная транскрипция происходит в сердцевине вириона. Многие вопросы, касающиеся цикла репродукции этого вируса, еще не выяснены.
Вирус гепатита В является представителем третьей группы. Двунитиевая кольцевая ДНК этого вируса имеет укороченную на 1/3 плюс-цепь и поэтому сначала достраивается с помощью ДНК-полимеразы, включенной в вирион, и превращается в сверхспиральную молекулу. Эта молекула транскрибируется с образованием молекул РНК двух классов-мРНК, кодирующей белки, и геномной РНК, которую затем транскрибирует обратная транскриптаза, синтезируя геномную ДНК.
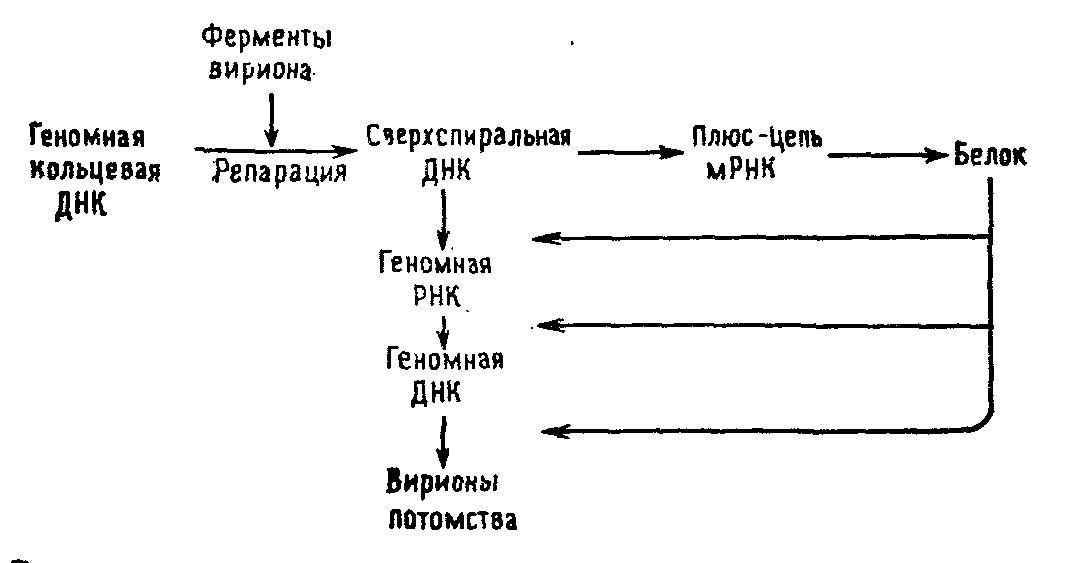
Размножение вируса гепатита В.
Парвовирусы и цирциновирусы входят в четвертуюю группу. Их геном представлен линейной или кольцевой плюс – или минус нитью ДНК. Для их размножения требуются синтез в ядре клетки ДНК-цепи, комплементарной одноцепочечной геномной ДНК, и последующая транскрипция генома.
Трансляция вирусных белков
Стратегия вирусного генома, использующего клеточный аппарат трансляции, должна быть направлена на создание механизма для подавления трансляции собственных клеточных иРНК и для избирательной трансляции вирусных иРНК, которые всегда находятся в значительно меньшем количестве, чем клеточные матрицы. Этот механизм реализуется на уровне специфического узнавания малой рибосомальной субъединицей вирусных иРНК, т. е. на уровне формирования инициирующего комплекса. Поскольку многие вирусы не подавляют синтез клеточных иРНК, в зараженных клетках возникает парадоксальная ситуация: прекращается трансляция огромного фонда функционально активных клеточных иРНК, и на освободившихся рибосомах начинается трансляция одиночных молекул вирусных иРНК. Специфическое узнавание рибосомой вирусных иРНК осуществляется за счет вирусспецифических инициаторных факторов.
Два способа формирования вирусных белков:
иРНК транслируется в гигантский полипептид-предшественник, который после синтеза последовательно нарезается на зрелые функционально активные белки; 2) иРНК транслируется с образованием зрелых белков, или белков, которые лишь незначительно модифицируются после синтеза.
иРНК транслируется с образованием зрелых белков, или белков, которые лишь незначительно модифицируются после синтеза.
Первый способ трансляции характерен для РНК-со-держащих «плюс-нитевых» вирусов — пикорнавирусов и тогавирусов. Их иРНК транслируется в гигантскую поли пептидную цепь, так называемый полипротеид, который сползает в виде непрерывной ленты с рибосомного «конвейера» и нарезается на индивидуальные белки нужного размера. Нарезание вирусных белков является многоступенчатым процессом, осуществляемым как вирусспецифическими, так и клеточными протеазами.
Второй способ формирования белков характерен для ДНК-содержащих вирусов и большинства РНК-содержащих вирусов. При этом способе синтезируются короткие моноцистронные иРНК в результате избирательной транскрипции одного участка генома (гена).
Вирусные белки в процессе инфекции синтезируются в избыточном количестве. Так, только 35% всех вирусиндуцированных белков герпесвирусов включается в вирионы. Вирусные белки накапливаются и в ядре и в цитоплазме, однако это не говорит о месте их синтеза.
СБОРКА ВИРУСНЫХ ЧАСТИЦ
Синтез компонентов вирусных частиц в клетке разобщен и может протекать в разных структурах ядра и цитоплазмы. При таком дисъюнктивном способе репродукции образование вирусной частицы возможно лишь в том случае, если вирусные нуклеиновые кислоты и белки обладают способностью при достаточной концентрации узнавать друг друга в многообразии клеточных белков и нуклеиновых кислот и самопроизвольно соединяться друг с другом, т. е. способны к самосборке. В основе самосборки лежит специфическое белокнук-леиновое и белок-белковое узнавание, которое может происходить в результате гидрофобных, солевых и водородных связей, а также стерического соответствия. Белокнуклеиновое узнавание ограничено небольшим участком молекулы нуклеиновой кислоты и определяется уникальными последовательностями нуклеотидов в неко дирующей части вирусного генома. С этого узнавания участка генома вирусными капсидными белками начинается процесс сборки вирусной частицы. Присоединение остальных белковых молекул осуществляется за счет специфиче-ческих белок-белковых взаимодействий или неспецифических белокнуклеиновых взаимодействий.
В связи с разнообразием структуры вирусов животных разнообразны и способы формирования вирионов, однако можно сформулировать следующие общие принципы сборки.
У просто устроенных вирусов формируются прови-рионы, которые затем в результате модификаций белков превращаются в вирионы. У сложно устроенных вирусов сборка осуществляется многоступенчато. Сначала формируются нуклеокапсиды или сердцевины, с которыми взаимодействуют белки наружных оболочек.
Сборка сложно устроенных вирусов (за исключением сборки вирусов оспы и реовирусов) осуществляется на клеточных мембранах. Сборка ядерных вирусов происходит с участием ядерных мембран, сборка цитоплазмати-ческих вирусов — с участием мембран эндоплазматическои сети или плазматической мембраны, куда независимо друг от друга прибывают все компоненты вирусной частицы.
У ряда сложно устроенных вирусов существуют специальные гидрофобные белки, выполняющие функции посредников между сформированными нуклеокапсидами и вирусными оболочками. Такими белками являются матриксные белки у ряда «минус-нитевых» вирусов (ортомиксовирусов, парамиксовирусов, рабдовирусов).
Сборка нуклеокапсидов, сердцевин, провирионов и вирионов происходит не во внутриклеточной жидкости, а в специальных структурах, предсуществующих в клетке или индуцированных вирусом («фабриках»).
Сложно устроенные вирусы для построения своих частиц используют ряд элементов клетки-хозяина, например липиды, некоторые ферменты, у ДНК-геномного SV40 — гистоны, у оболочечных РНК-геномных вирусов — актин, а в составе ареновирусов обнаружены даже рибосомы. Клеточные молекулы несут определенные функции в вирусной частице, однако включение их в вирион может явиться и следствием случайной контаминации, как, например, включение ряда ферментов клеточных оболочек или клеточных нуклеиновых кислот.