

Содержание
Стр.
Введение………………………………………………………………………………….3
Белки и ферменты, участвующие в репликации ДНК………………………………...4
Репликация ДНК…………………………………………………………………………6
Инициация…………………………………………………………………………....6
Элонгация…………………………………………………………………………….7
Терминация…………………………………………………………………………..8
Заключение……………………………………………………………………………….8
Список использованных источников…………………………………………………...9
Введение
Генетическая программа клеточных организмов записана в нуклеотидной последовательности ДНК. Для сохранения уникальных свойств организма необходимо точное воспроизведение этой последовательности в каждом последующем поколении. Во время деления клетки содержание ДНК должно удвоиться, чтобы каждая дочерняя клетка могла получить полный спектр ДНК.
Процесс удвоения родительских молекул геномной ДНК во время воспроизводства клеток живого организма получил название репликации, или репликативного синтеза ДНК.
Процесс репликации состоит из трех стадий: инициации (начало процесса), элонгации (собственно синтез) и терминации (окончание процесса).
Репликация ДНК является примером матричного синтеза биологических макромолекул. Основу хромосомы составляет одна непрерывная двухцепочечная молекула ДНК. Во время репликации каждая из цепей родительской ДНК служит матрицей для синтеза комплементарной дочерней цепи. Положение каждого нуклеотида в строящейся цепи ДНК по правилам комплементарности (G-C и A-T) однозначно определяется положением соответствующего нуклеотида матрицы.
Механизм, происходящий при репликации ДНК химической реакции, заключается в переносе остатка дезоксирибонуклеозидмонофосфата от дезоксирибонуклеозидтрифосфата на концевой нуклеотидный остаток растущей в процессе синтеза нуклеотидной цепи (рис.1).
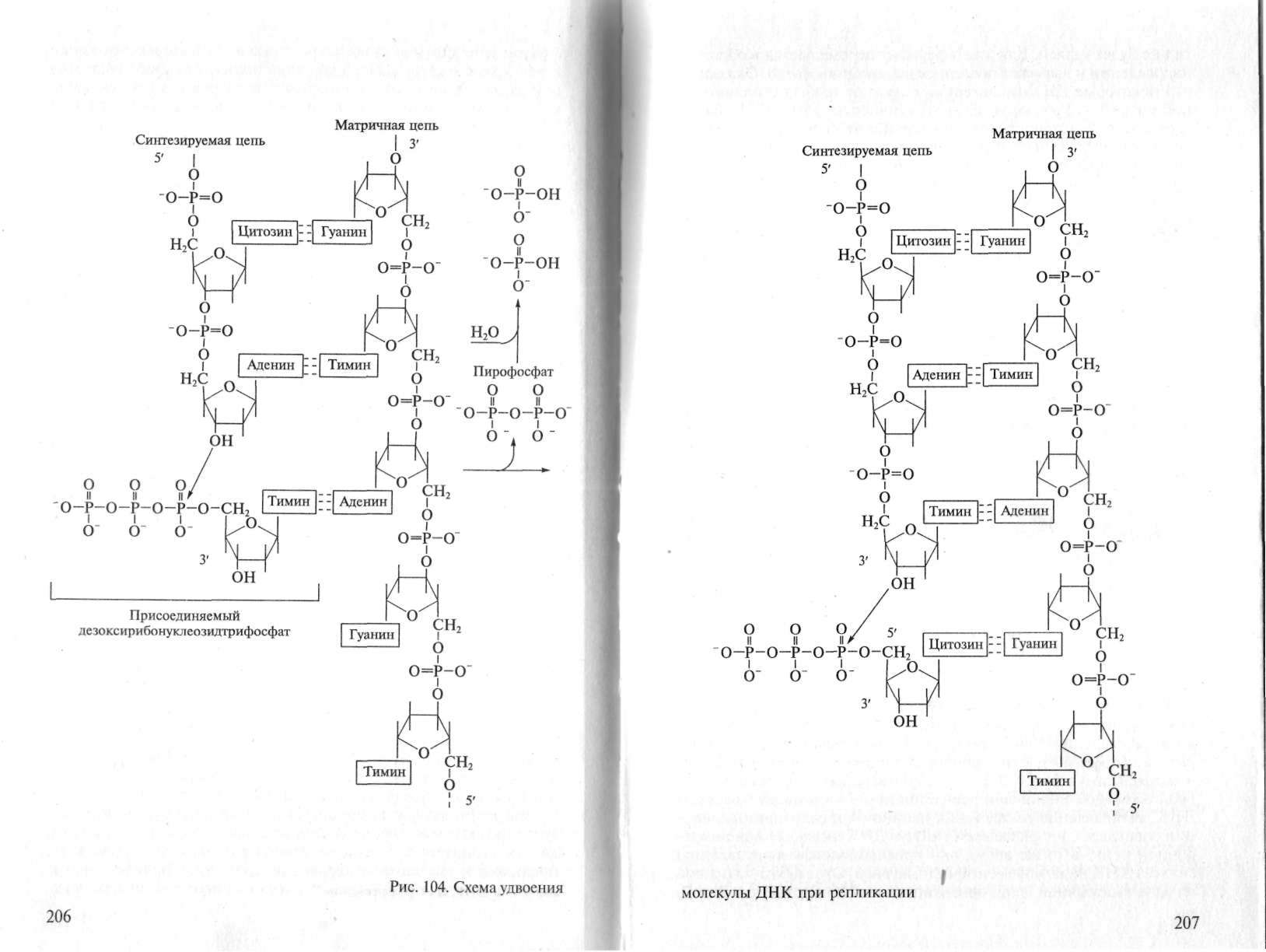
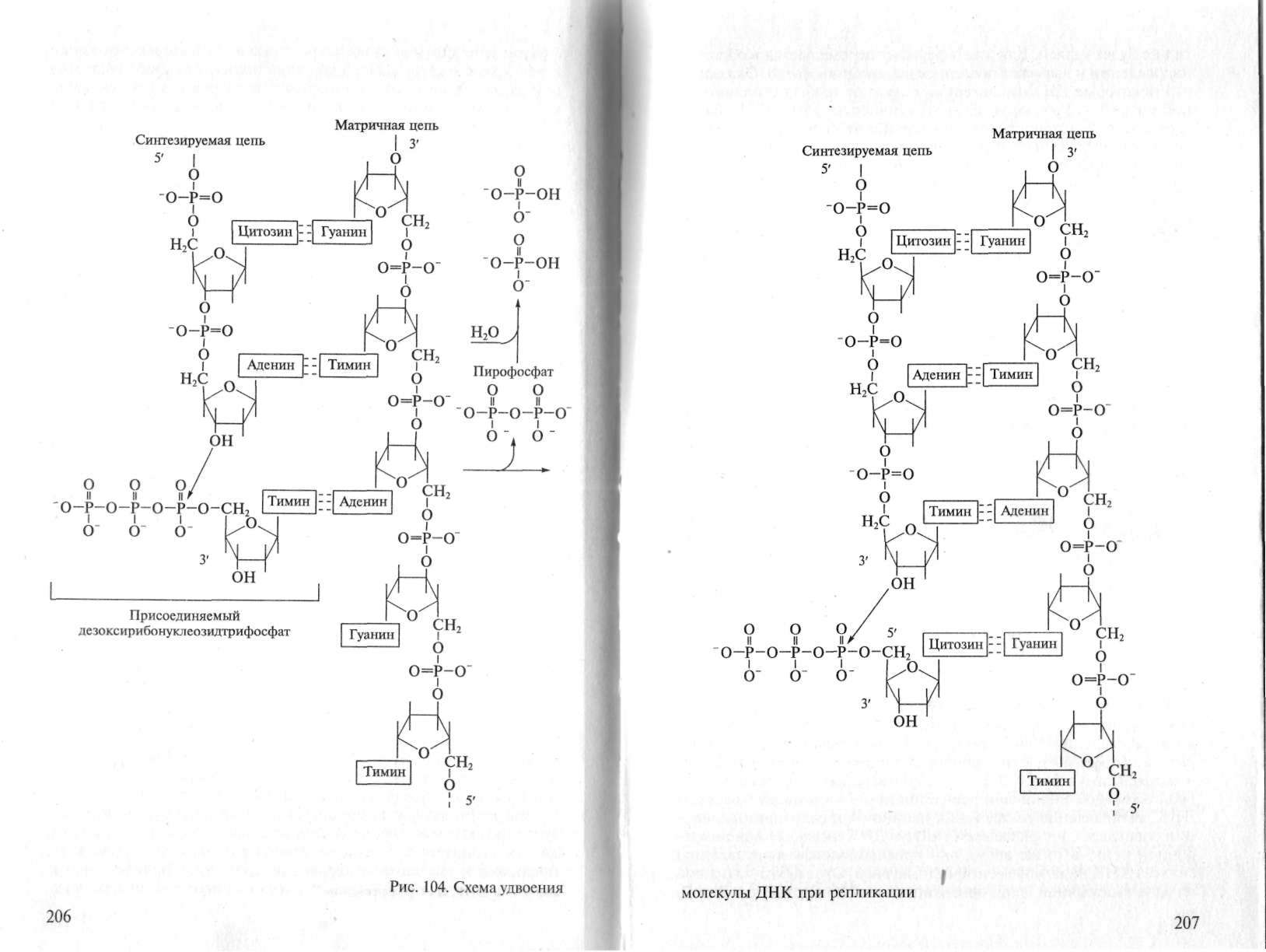
Рис.1. Схема удвоения молекулы ДНК при репликации
Белки и ферменты, участвующие в репликации днк
ДНК-полимеразы. Катализируют комплементарное копирование одноцепочечной матрицы ДНК.
Полимеризация нуклеотидов происходит в одном направлении от 5’ к 3’ концу новой цепи. И синтезируют цепь антипараллельной матричной цепи ДНК. Так как копирование ДНК требует высокой точности, то существует специальный механизм проверки точности копирования (механизм коррекции).
ДНК-полимеразы проверяют комплементарность каждого нуклеотида матрицы дважды:
1) перед включением его в состав растущей цепи;
2) перед тем, как включить следующий нуклеотид.
Если происходит ошибка, то неправильный нуклеотид удаляется. Для этого фермент перемещается в обратном направлении и вырезает последнее добавленное звено. Наряду с 5’→3’ эндонуклеотидной активностью некоторые ДНК-полимеразы обладают 3’→5’ экзонуклеазной активностью.
ДНК-праймаза. Не способны инициировать синтез новых цепей ДНК, они могут только добавлять дезоксирибонуклеотиды к 3’-концу уже имеющейся полинуклеотидной цепи. Для начала синтеза ДНК этим ферментам необходимы затравки-праймеры. В качестве праймеров выступают короткие олигодезоксирибонуклеиновые или олигорибонуклеотиды. На стадии инициации репликации короткую РНК-затравку синтезируют специфичные ферменты ДНК-праймаза. У бактерий этот фермент является самостоятельным белком, а у эукариот он входит в состав ДНК-полимеразы как субъединица. В любом случае праймаза – это фермент, отличный от РНК-полимераз, которые синтезируют разнообразные клеточные РНК. После того как будет синтезирован РНК-праймер, подключается ДНК-полимераза и продолжает наращивать цепь.
ДНК-лигаза. ДНК-лигазы вирусов, бактерий, млекопитающих соединяют 5’-фосфатную и 3’-гидроксильную группы нуклетидов, находящихся на противоположных концах одноцепочечного разрыва в дуплексе ДНК. В результате образуется фосфодиэфирная связь, ликвидирующая этот разрыв (рис.2).
ДНК-лигаза Е. coli — это одиночный полипептид. Для образования фосфодиэфирной связи между концами нуклеотидных цепей ДНК-лигазы используют энергию гидролиза АТР либо NAD. Реакция протекает в три стадии:
Реакция аденилирования ДНК-лигазы. Аденилильный остаток NAD или АТФ переносится на ε-аминогруппу лизинового остатка в активном центре лигазы с одновременным высвобождением пирофосфата (в случае АТФ) или никотин-амидмононуклеотида (в случае NAD).
Реакция трансаденилирования. Остаток адениловой кислоты с аденилатлигазы перебрасывается на свободную фосфатную группу 5’-углеродного атома рибозы концевого нуклеотида.
Реакция лигирования. Между сближенными активированной аденилатом 5’-фосфатной группой и 3’-ОН-группой фрагмента цепи ДНК происходит образование фосфодиэфирной связи с выделением АМР.
ДНК-хеликаза. Раскручивание, или расплетание, спирали происходит в локальном участке ДНК. Эту реакцию осуществляет хеликаза — ДНК-зависимая АТРаза, использующая энергию гидролиза АТР для расплетания двойной спирали ДНК. Хеликазы имеют кольцевую (тороидальную) структуру, образованную шестью субъединицами. Такие гексамерные хеликазы кольцеобразной формы обнаружены у фагов, вирусов, бактерий, архей, эукариот (рис. 3). Хеликаза, движимая гидролизом АТР, однонаправленно перемещается по одной из цепей ДНК (вероятно, за счет ее конформационных изменений), расплетая перед собой двойную спираль, в результате чего возникает вилка (Y) из двуцепочечного участка ДНК и двух одноцепочечных ветвей (рис.3).

Рис.2. Механизм действия ДНК-лигазы: Риб – рибоза; А – аденин; РР - пирофосфат
|
|
Рис.3. ДНК-хеликазы:
А – четвертичная структура гексамерных ДНК-хеликаз: фага Т4 (1),вирусаSV(2),DnaBE. Coli (3); Б – схема расплетения двойной спирали ДНК-хеликазой |

Белки, связывающиеся с одноцепочечной ДНК, или SSB-белки. SSB-белки связываются с сахарофосфатным остовом одиночных цепей ДНК, не закрывая оснований, что не мешает комплементарному присоединению нуклеотидов в ходе репликации. SSB-белок Е. coli наиболее изучен, он представляет собой тетрамер, характеризуется высокой степенью асимметрии молекулы. SSB-белки стабилизируют одноцепочечную ДНК, обеспечивая условия для комплементарного спаривания, удаляют Возможные элементы вторичной структуры ДНК(например, предотвращают образование шпилечных структур); связывание одноцепочечной ДНК с SSB-белками стимулирует ДНК-полимеразу и повышает точность ее работы. У эукариот таковым белком является ядерный репликативный белок A (RPA), представляющий гетеротример.