

BIO_-_vsyo
.pdfПрофаза II. Не отличается существенно от профазы митоза. Хромосомы видны в световой микроскоп в виде тонких нитей.
Метафаза II. В каждой из дочерних клеток формируется веретено деления. Хромосомы располагаются в экваториальных плоскостях гаплоидных клеток независимо друг от друга. Эти экваториальные плоскости могут лежать в одной плоскости, могут быть параллельны друг другу или взаимно перпендикулярны.
Анафаза II. Хромосомы разделяются на хроматиды (как при митозе). Получившиеся однохроматидные хромосомы в составе анафазных групп перемещаются к полюсам клеток.
Телофаза II. Однохроматидные хромосомы полностью переместились к полюсам клетки, формируются ядра. Содержание ДНК в каждой из клеток становится минимальным и составляет 1с.
67. Гаметогенез и его этапы. Сравнение овогенеза и сперматогенеза.
Гаметогенез — это процесс образования половых клеток. Протекает он в половых железах (в яичниках у самок и в семенниках у самцов). Гаметогенез в организме женской особи сводится к образованию женских половых клеток (яйцеклеток) и носит название овогенеза. У особей мужского пола возникают мужские половые клетки (сперматозоиды), процесс образования которых называется сперматогенезом.
Стадии гаметогенеза 1. Стадия размножения.
Клетки, из которых в последующем образуются мужские и женские гаметы, называются сперматогониями и овогониями соответственно.
Они несут диплоидный набор хромосом 2n2c.
На этой стадии первичные половые клетки многократно делятся митозом, в результате чего их количество существенно возрастает.
Сперматогонии размножаются в течение всего репродуктивного периода в мужском организме. Размножение овогоний происходит главным образом в эмбриональном периоде.
2. Стадия роста.
Kлетки увеличиваются в размерах(за счет увеличения объема цитоплазмы) и превращаются в сперматоциты и овоциты I порядка (последние достигают особенно
больших размеров в связи с накоплением питательных веществ в виде желтка и белковых гранул).
Эта стадия соответствует интерфазе I мейоза.
Важное событие этого периода — репликация молекул ДНК при неизменном количестве хромосом. Они приобретают двунитчатую структуру: генетическая формула клеток в этот период выглядит как 2n4c.
3. Стадия созревания.
Происходят два последовательных деления — редукционное (мейоз I) и эквационное (мейоз II), которые вместе составляют мейоз.
После первого деления (мейоза I) образуются сперматоциты и овоциты II порядка (с генетической формулой n2c), после второго деления (мейоза II) — сперматиды и зрелые яйцеклетки (с формулой nc) с тремя редукционными тельцами, которые погибают и в процессе размножения не участвуют.
Таким образом, в результате стадии созревания один сперматоцит I порядка (с формулой 2n4c) дает четыре сперматиды (с формулой nc), а один овоцит I порядка (с формулой 2n4c) образует одну зрелую яйцеклетку (с формулой nc) и три редукционных тельца.
4. Стадия формирования, или спермиогенеза (только при сперматогенезе).
В результате этого процесса каждая незрелая сперматида превращается в зрелый сперматозоид (с формулой nc), приобретая все структуры, ему свойственные. Ядро сперматиды уплотняется, происходит сверхспирализация хромосом, которые
становятся функционально инертными. Комплекс Гольджи перемещается к одному из полюсов ядра, формируя акросому. К другому полюсу ядра устремляются центриоли, причем одна из них принимает участие в формировании жгутика. Вокруг жгутика спирально закручивается одна митохондрия. Почти вся цитоплазма сперматиды отторгается, поэтому головка сперматозоида ее почти не содержит.
Отмеченные выше различия в ходе овогенеза и сперматогенеза имеют определенный биологический смысл, связанный с разным функциональным назначением мужских и женских гамет (помимо переноса генетической информации). Накопление в цитоплазме яйцеклетки большого количества запасных питательных веществ необходимо, так как на этой «базе» осуществляется развитие дочернего организма из оплодотворенного яйца. Неравномерное клеточное деление при овогенезе и обеспечивает формирование крупной яйцеклетки. Функция же сперматозоидов заключается в отыскании яйцеклетки, проникновении в нее и доставке своего хромосомного набора. Их существование кратковременно, а поэтому нет необходимости в запасании большого количества веществ в цитоплазме. А поскольку сперматозоиды в массе гибнут в процессе поиска яйцеклетки, их образуется огромное количество.
Центральное событие в процессе гаметогенеза — редукция диплоидного набора хромосом (в ходе мейоза) и формирование гаплоидных гамет.
68. Закономерности сперматогенеза у млекопитающих и человека.(схема
67)
Последовательные стадии развития сперматозоидов получили название сперматогенеза.
1. Стадия размножения.
Клетки, из которых в последующем образуются мужские гаметы, называются сперматогониями. Они несут диплоидный набор хромосом 2n2c.
На этой стадии первичные половые клетки многократно делятся митозом, в результате чего их количество существенно возрастает.
Сперматогонии размножаются в течение всего репродуктивного периода в мужском организме.
2. Стадия роста.
Kлетки увеличиваются в размерах (за счет увеличения объема цитоплазмы) и превращаются в сперматоциты I порядка Эта стадия соответствует интерфазе I мейоза.
Важное событие этого периода — репликация молекул ДНК при неизменном количестве хромосом. Они приобретают двунитчатую структуру: генетическая формула клеток в этот период выглядит как 2n4c.
3. Стадия созревания.
Происходят два последовательных деления — редукционное (мейоз I) и эквационное (мейоз II), которые вместе составляют мейоз.
После первого деления (мейоза I) образуются сперматоциты II порядка (с генетической формулой n2c), после второго деления (мейоза II) — сперматиды (с формулой nc) . Таким образом, в результате стадии созревания один сперматоцит I порядка (с формулой 2n4c) дает четыре сперматиды (с формулой nc),
4. Стадия формирования, или спермиогенеза (только при сперматогенезе).
Конечный этап в сперматогенезе связан с дифференциацией, которая заканчивается тем, что каждый из сравнительно больших, сферических неподвижных сперматидов превращается в небольшой вытянутый подвижный сперматозоид (с формулой nc). Ядро сперматиды уплотняется, происходит сверхспирализация хромосом, которые становятся функционально инертными. Комплекс Гольджи перемещается к одному из полюсов ядра, формируя акросому. К другому полюсу ядра устремляются центриоли, причем одна из них принимает участие в формировании жгутика. Вокруг жгутика спирально закручивается одна митохондрия. Почти вся цитоплазма сперматиды отторгается, поэтому головка сперматозоида ее почти не содержит.
Каждый сперматозоид человека состоит из трех отделов — головки, средней части и хвоста. В головке сперматозоида располагается ядро. В нем содержится гаплоидный набор хромосом. Головка снабжена акросомой, которая содержит литические ферменты, необходимые сперматозоиду для вступления в яйцеклетку. В головке локализуется также две центриоли — проксимальная, которая побуждает деление оплодотворенной сперматозоидом яйцеклетки, и дистальная, которая дает начало аксимальному стержню хвоста. В средней части сперматозоида располагаются базальное тельце хвоста и митохондрии. Хвост (отросток) сперматозоида образован внутренним аксимальным стержнем и внешним футляром, имеющим цитоплазматическое происхождение. Сперматозоиды человека характеризуются значительной подвижностью.
Различия в ходе овогенеза и сперматогенеза имеют определенный биологический смысл, связанный с разным функциональным назначением мужских и женских гамет (помимо переноса генетической информации). Накопление в цитоплазме яйцеклетки большого количества запасных питательных веществ необходимо, так как на этой «базе» осуществляется развитие дочернего организма из оплодотворенного яйца. Неравномерное клеточное деление при овогенезе и обеспечивает формирование крупной яйцеклетки. Функция же сперматозоидов заключается в отыскании яйцеклетки, проникновении в нее и доставке своего хромосомного набора. Их существование кратковременно, а поэтому нет необходимости в запасании большого количества веществ в цитоплазме. А поскольку сперматозоиды в массе гибнут в процессе поиска яйцеклетки, их образуется огромное количество.
69. Закономерности овогенеза у млекопитающих и человека. (Схема.67)
Гаметогенез в организме женской особи сводится к образованию женских половых клеток (яйцеклеток) и носит название овогенеза. Он протекает он в половых железах (в яичниках у самок). Его функциями являются обеспечение гаплоидного набора хромосом в ядре яйцеклетки и обеспечение питательных потребностей зиготы
Стадии овогенеза
1.Стадия размножения.
Первичные половые клетки многократно делятся митозом, в результате чего их количество существенно возрастает. Размножение оогоний происходит главным образом в эмбриональном периоде.
2. Стадия роста.
Kлетки увеличиваются в размерах (за счет увеличения объема цитоплазмы) и превращаются в овоциты I порядка. Эта стадия соответствует интерфазе I мейоза.
Важное событие этого периода — репликация молекул ДНК при неизменном количестве хромосом. Они приобретают двунитчатую структуру: генетическая формула клеток в этот период выглядит как 2n4c.
Втечение дифференциации яйцеклеток формируются мембраны, уменьшается в размере их ядро.
3. Стадия созревания.
Происходят два последовательных деления — редукционное (мейоз I) и эквационное (мейоз II), которые вместе составляют мейоз. Ко времени рождения девочки все первичные ооциты уже находятся в профазе первого мейотического деления.
Ооцит в состоянии покоя окружен одним слоем уплощенных фолликулярных клеток и образует вместе с ними примордиальный фолликул. Ооциты, не окруженные фолликулами, дегенерируют. Некоторые из фолликулов периодически начинают развитие-фолликулогенез. Сначала клетки фолликула делятся, образуя многослойную оболочку ооцита; сам ооцит растет, вокруг него образуется прозрачная многослойная зона-формируется развивающийся фолликул. Затем в некоторых фолликулах образуется зона-формируется развивающийся фолликул. Затем в некоторых фолликулах образуется наполненная жидкостью полость-антриум, и развивающийся фолликул превращается в антральный. Внутри фолликула ооцит постепенно окружается клеточной массой яйценосного бугорка. До достижении половой зрелости процессы развития фолликулов завершаются дегенерацией. С наступлением половой зрелости также одновременно развиваются многие фолликулы, однако лишь некоторые высвобождают находящиеся в них ооциты в процессе овуляции. Большая часть фолликулов дегенерирует на той или иной стадии. Продолжительность роста фолликула составляет приблизительно 6 месяцев.
Всоответствии с менструальным циклом раз в месяц происходит повышение уровня лютеинизирующего гормона, выделяемого гипофизом, что побуждает примерно 20 антральных фолликулов к ускоренному росту. Однако завершает созревание и претерпевает овуляцию лишь один из фолликулов. Ооцит первого порядка, находящийся в этом фолликуле, завершает первое деление мейоза, образуя полярное тельце и превращаясь в ооцит второго порядка. Ооцт второго порядка, развитие которого останавливается на стадии метафазы второго деления мейоза, освобождается вместе с полярным тельцем и часть окружающих его клеток в момент разрыва фолликула на поверхности яичника. Ооцит второго порядка завершает второе мейотическое деление только в том случае, если он будет оплодотворен. После овуляции опустевший фолликул превращается в эндокринное образование-желтое тело. Оно секретирует прогестерог, готовя матку к приему оплодотворенного ооцита. Если оплодотворение не происходит, желтое тело деградирует, а слизистая оболочка матки отслаивается и выводится из организма во время менструации.
Отличительные особенности
Питательный материал (желток) первичного ооцита не распределяется поровну между четырьмя клетками.. Основное количество желтка сохраняется в одной большой клетке. Первые и вторые полярные тельца получают в результате делений те же самые хромосомные наборы, что и вторичные ооциты, но они не становятся половыми клетками. Яйцеклетки млекопитающих имеют овальную или несколько вытянутую форму и характеризуются типичными чертами клеточного строения. Они содержат все структуры, характерные для соматических клеток. Одна из характерных особенностей яйцеклеток заключается в сложности строения их оболочек. Различают первичную, вторичную и третичную оболочки яйцеклеток. Первичная оболочка (внутренняя) формируется на стадии ооцита и представляет собой поверхностный слой ооцита. Вторичная (средняя) оболочка полностью сформирована фолликулярными клетками, а третичная (наружная) образована веществами, представляющими собой продукты секреции желез яйцеводов, через которые проходят яйцеклетки.
Различия в ходе овогенеза и сперматогенеза имеют определенный биологический смысл,
связанный с разным функциональным назначением мужских и женских гамет (помимо переноса генетической информации). Накопление в цитоплазме яйцеклетки большого
количества запасных питательных веществ необходимо, так как на этой «базе» осуществляется развитие дочернего организма из оплодотворенного яйца. Неравномерное клеточное деление при овогенезе и обеспечивает формирование крупной яйцеклетки. Функция же сперматозоидов заключается в отыскании яйцеклетки, проникновении в нее и доставке своего хромосомного набора. Их существование кратковременно, а поэтому нет необходимости в запасании большого количества веществ в цитоплазме. А поскольку сперматозоиды в массе гибнут в процессе поиска яйцеклетки, их образуется огромное количество.
70. Оплодотворение, его формы и биологическая функция. Моно- и полиспермия.
Оплодотворение-это процесс объединения мужской и женской гамет, который приводит к формированию зиготы и последующему развитию нового организма. В процессе оплодотворения происходит установление диплоидного набора хромосом в зиготе, что определяет выдающееся биологическое значение этого процесса.
Взависимости от видовой принадлежности организмов у животных, размножающихся половым путем, различают наружное и внутреннее оплодотворение.
Наружное оплодотворение происходит в окружающей среде, в которую поступают мужские и женские половые клетки. Например, наружным является оплодотворение у рыб. Выделяемые ими мужские (молока) и женские (икра) половые клетки поступают в воду, где и происходит их «встреча» и объединение.
Внутреннее оплодотворение обеспечивается переносом сперматозоидов из мужского организма в женский в результате полового акта. Такое оплодотворение встречается у млекопитающих, причем центральным моментом здесь является исход встречи между половыми клетками. Считают, что в яйцеклетку этих животных проникает ядерное содержимое лишь одного сперматозоида. Что касается цитоплазмы сперматозоида, то у одних животных она поступает в яйцеклетку в небольшом количестве, у других совсем не поступает в яйцеклетку.
У человека оплодотворение происходит в верхней части фаллопиевой трубы, причем в оплодотворении, как и у других млекопитающих, участвует лишь один сперматозоид, ядерное содержимое которого поступает в яйцеклетку. Иногда в фаллопиевой трубе может оказаться не одна, а две или более яйцеклеток, в результате чего возможно рождение двоен, троен и т. д.
Врезультате оплодотворения в оплодотворенной яйцеклетке восстанавливается диплоидный набор хромосом. Яйцеклетки способны к оплодотворению в течение примерно 24 часов после овуляции, тогда как оплодотворяющая способность сперматозоидов сохраняется до 48 часов.
Взависимости от количества особей, принимающих участие в половом размножении, различают: -перекрестное оплодотворение-оплодотворение, в котором принимают участие гаметы, образованные разными организмами -самооплодотворение-оплодотворение, при котором сливаются гаметы, образованные одним и тем же организмом (ленточные черви).
Существует два типа оплодотворения: моно- и полиспермия.
При моноспермии благодаря особым механизмам в яйцеклетку проникает лишь один сперматозоид. Этот вид оплодотворения широко распространен у животных с наружным осеменением, хотя свойствен также и млекопитающим.
При полиспермии в яйцеклетку проникает до нескольких десятков сперматозоидов, однако ядро только одного из них соединяется с женским пронуклеусом, остальные посредством специальных механизмов исключаются из развития. Этот тип оплодотворения присущ животным с внутренним осеменением (членистоногие, моллюски, хордовые). Оболочка яйцеклетки насекомых и многих рыб содержит отверстия, через которые сперматозоиды проникают в яйцеклетку. У млекопитающих такие отверстия отсутствуют, и, чтобы достигнуть ооплазмы, сперматозоиды должны пройти через толщу оболочек. Для этого у сперматозоида имеются специальные органеллы-акросомы, располагающиеся на вершине его головки. Погрузившись в ооплазму, головка сперматозоида ориентируется в глубь яйцеклетки и претерпевает сложные изменения. При этом его ядро постепенно преобразуется в мужской пронуклеус. Эти преобразования идут одновременно с формированием женского пронуклеуса. Мужской и женский пронуклеусы сближаются в центре яйцеклетки и затем сливаются, образуя единое ядро зиготы. На этом
завершается процесс оплодотворения. Образовавшаяся после оплодотворения зигота постепенно продвигается по направлению к матке и выходит в нее через несколько суток.
71. Морфологические и функциональные особенности зрелых гамет млекопитающих и человека
Гаметы - репродуктивные клетки, имеющ гаплоидный (одинарный) набор хром и уч в гаметном, половом размножении. При слиянии двух гамет обр зигота, развив в особь (или группу особей) с наследственными признаками обоих родительских организмов, продуцировших гаметы.
У некоторых видов возможно и развитие в организм одиночной гаметы (неоплодотворённой яйцеклетки) — партеногенез.
Морфология гамет и типы гаметогамии
Продуцируемые гаметы могут отличаться как по хромосомному набору (при гетерогаметности вида), величине и подвижности (способности к самостоятельному передвижению).
Если сливающиеся гаметы морфолог не отличаются друг от друга величиной, строением и хромосомным набором-это изогаметы(бесполые гам). Они подвижны, могут нести жгутики или быть амёбовидными. Изогамия типична для многих водорослей.
Анизогамия (гетерогамия)
Гаметы, способные к слиянию, различаются по размерам, подвижные микрогаметы несут жгутики, макрогаметы могут быть как подвижны (многие водоросли), так и неподвижны (лишённые жгутиков макрогаметы многих протистов).
Оогамия Сперматозоид и яйцеклетка.
Способные к слиянию гаметы одного биологического вида резко различаются по размерам и подвижности на два типа: мужские гаметы малого размера и крупные неподвижные женские гаметы — яйцеклетки. Различие размера гамет обусловлено тем,
что яйцеклетки содержат запас питательных веществ, достаточный для обеспечения нескольких первых делений зиготы при её развитии в зародыш.
Мужские гаметы — сперматозоиды животных и многих растений подвижны и обычно несут один или несколько жгутиков, исключением являются лишённные жгутиков мужские гаметы семенных растений — спермии, кот дост-ся к яйцеклетке при прорастании пыльцевой трубки, безжгутиковые сперматозоиды (спермии) нематод и членистоногих.
Хотя сперматозоиды несут митох, при оогамии от мужской гаметы к зиготе переходит ядерная ДНК, мтх ДНК наследуется зиготой только от яйцеклетки.
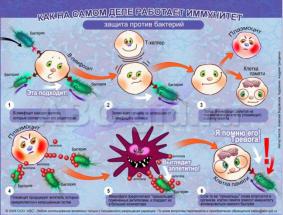
72. Клеточные факторы иммунной системы.
-лимфоциты(Т, B, NK) -моноциты -макрофаги -нейтрофилы -эозинофилы
-базофилы(тучные) клетки
Клеточн факторы врожденного иммунитета: Нейтрофилы и Макрофаги(моноциты)- «профессиональные» фагоциты.
Для уничтожения:
1.Кислороднонезависимый мех-м(гидролитические ферм)
2.Кислородзависимый(«кислородн» взрыв с обр н2о2 и аниона о2-, гипохлорида-для киллинга)
3.NO (и производные) после дополн активац с помощью Т-хелперов.
Продукция и секреция:

+Вещ-ва для уничтожения прод вне клетки +Сигнальные вещ-ва (цитокинов и тд…)
МОНОЦИТЫ и МАКРОФАГИ – Антигенпредставл кл (АПК) Представляют антигены-презентац антигенов в компл с МНС (главн комп гистосовместим) Т-клетками.
ДЕНДРИТНЫЕ кл-с отростками.
-захват, затем процессинг антигена
-презентац антигенов Т-лимфоцитам (гл АПК)
-костимулирующие сигналы Т-лимф
-др. функц-регуляц созревания Т-лимф в тимусе
БАЗОФИЛЫ(тучн кл) – Синтез и секреция медиаторов воспаления(гистамин, простагландин, гепарин…) Вызыв аллергич реакц, несут на поверхн рецепторы к иммуноглобулинам Е и Fe
ЭОЗИНОФИЛЫ – антителзависимое цитотоксическое вещ-во(действ в отношении личинок гельминтов) Уч в аллергич реакции, несут рецепторы к иммуноглобулинам Е и
Fe
NK-клетки (норм киллеры)-лимфоциты. Уничтожают кл опухолей и инфицированные вирусами. Цитотоксич действие-без апоптоза в отлич от Т и В лимфоцитов. Не имеют антиген спецефич рецепторов. Убивают кл, лишившиеся MHCl. Проявл антителзависимую кл цитотоксичность (связ «хвост» антитела с помощбю Fe рец и убивают кл с кот это антитело взаимодействует).
73. Т-лимфоциты – клеточные факторы приобретенного клеточного иммунитета.
Термин клеточный иммунитет используется для описания такого противоинфекционного или противоопухолевого иммунного ответа, в котором антителам принадлежит не ведущая, а вспомогательная роль.
Основу клеточного иммунитета составляют лимфоциты, которые для своего созревания переселяются из костного мозга в другой центральный орган лимфоидной системы - тимус (вилочковая железа). Эта ветвь лимфоцитов получила название тимус-зависимые, или Т-лимфоциты.
Т-лимфоциты выполняют следующие функции:
– являются основными эффекторами клеточного иммунитета (эти клетки опосредуют клеточные цитотоксические реакции, а также реакции ГЗТ);
–являются регуляторами воспаления, иммунных реакций и гемопоэза;
–участвуют в процессах репаративной и физиологической регенерации различных тканей. В организме человека Т-лимфоциты многократно покидают лимфоидные органы, попадая сначала в лимфу, затем в кровь, а из крови снова возвращаются в органы. За свою жизнь лимфоцит может проходить более 100 километров. Благодаря интенсивной циркуляции, лимфоциты, когда в них возникает потребность, быстро появляются в "горячих точках".
В тимусе формируются разные виды Т-клеток.
1.некоторые Т-лимфоциты обладают способностью разрушать клетки, содержащие чужеродный антиген, их назвали цитотоксическими или "киллерами".
2.другая разновидность лимфоцитов - Т-хелперы - первыми распознают чужеродные вещества. Т-хелперы не способны вырабатывать антитела и убивать клетки-мишени, но, распознавая чужеродный антиген, они реагируют на него выработкой различных факторов, которые необходимы для размножения и созревания В-клеток и Т-киллеров. Именно Т-хелперы распознают антигены и влияют на деятельность других типов Т- клеток, оказывают помощь В-клеткам в образовании антител. По их командам иммунная система направляет Т-лимфоцитов-киллеров, задача которых убивать зараженные клетки. 3.существуют еще Т-супрессоры, которые подавляют активность иммунного ответа, когда необходимость в нем отпадает. Если иммунные клетки будут продолжать работать, то
будут поражаться собственные здоровые клетки организма, что приведет к развитию различных болезней (их называют аутоиммунными).
74. В-лимфоциты – клеточные факторы приобретенного гуморального иммунитета.
Приобретенный иммунитет:
•Т и В лимфоц
•MHC (у чел HLA)
•Антитела
•Клеточный и Гуморальный иммунный ответ
В-лимфоциты-гуморальный иммун. Распознание нерасщепл антигенов. Наивные В-лимф. Плазматические В-кл памяти-вторичный иммунитет.
Отличия Т- и В-лимфоцитов:
-по месту антигеннезависим дифференц: Т-лимфоциты в тимусе, В-лимфоциты в лимфоидной ткани других органов.
-по выполняемым функциям: Т-лимфоциты обеспечивают преимущественно клеточный иммунитет, В-лимфоциты – гуморальный иммунитет. Функционально Т- и В-клетки делят
на субпопуляции. Среди Т-лимфоцитов выделяют Тх (хелперы) – активируют эффекторные клетки, Тк (киллеры) – эффекторные цитотоксические клетки, Тс (супрессоры) – подавляют иммунный ответ, Т-лимфоциты памяти.
В-лимфоциты дифференцируются в плазматические клетки, вырабатывающие иммуноглобулины (антитела) и в клетки памяти, несущие информацию о встрече с какимлибо антигеном.
-по содержанию в крови: Т-лимфоцитов – 70-80%, В-лимфоцитов — 10-20%.
Функция лимфоцитов:
1) участие и обеспечение иммунных реакций.
В-лимфоциты проходят предварительную обработку в печени и костном мозге. Существуют два важных различия между В- и Т-лимфоцитами.
- Во-первых, В-лимфоциты активно секретируют реактивные агенты, называемые антителами, в отличие от Т-лимфоцитов, реагирующих с антигеном непосредственно. Антитела — это крупные белковые молекулы, способные соединяться с антигенной субстанцией и разрушать ее.
- Во-вторых, разнообразие В-лимфоцитов выражено больше, чем у Т-лимфоцитов, т.е. формируются миллионы типов В-лимфоцитарных антител с разными специфическими реактивностями.

РАЗДЕЛ 2
1. Уровни организации генетического аппарата клетки (геном, генотип, кариотип).
ГЕНОМ-общ протяж-ть ДНК в гаплоидном наборе хр-м. Термин «геном» был предлож Г. Винклером в 1920 г. для опис-я совокуп-ти генов, заключ-х в гапл-м наборе хр-м орг-в одного биологич-го вида. Первонач смысл этого термина указ на то, что понятие генома в отличие от генотипа явл генетич-й характер-кой вида в целом, а не отдел-й особи. С развем молекул-й генетики знач-е данного термина измен-сь. Генотип — совокупн-ть генов данного орг-ма, к-ая, в отличие от понятий генома и генофонда, характериз особь, а не вид. Вместе с факторами внешн среды генотип определ фенотип орг-ма. Фенотип и генотип различ-ся: 1- по ист-ку инф-ции (генотип определ-ся при изучении ДНК особи, фенотип регистрир-ся при наблюдении внешн вида орг-ма). 2- Генотип не всегда соотв одному и тому же фенотипу.Нек-ые гены проявл-ся в фенотипе только в определ-х усл-ях. Кариотип –совок-ть призн-в (число, размеры, форма и т. д.) полного набора хр-м, присущая кл-кам данного биологич-го вида (видовой кариотип), данного организма
(индивидуальный кариотип) или линии (клона) клеток.
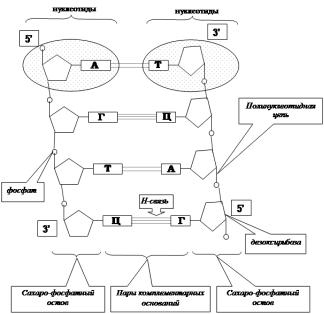
2. Структура ДНК. Модель Дж. Уотсона и Ф. Крика.
ДНК – дезоксирибонуклеин к-та – биологич макромолекула, носитель генетич-й инф-ции во всех эукариотич-х кл-ках. Трехмерн модель пространств-го строения двухцепочечной ДНК была описана в 1953 г. Дж. Уотсоном и Френсисом Криком. Согласно этой модели мол-ла ДНК сост из двух полинуклеотидных цепей, к-рые образ правую спираль (винтовую линию) относительно одной и той же оси. Направл-е цепей взаимно противоположное. Структура ДНК – полимер, структурной единицей которого является нуклеотид (сост из: азот-го основ-я: пуринового – аденин (А) или гуанин (Г) или пиримидинового – цитозин (Ц) или тимин (Т); углевода дезоксирибозы (пятиуглер-е сахарное кольцо); остатка фосф-ой к-ты(НРО3*). Сахарофосфатный остов располаг по периферии двойной спирали, а азотистые основ-я наход-ся внутри и их плоскости перпендикулярны оси спирали. Между основ-ями образ-ся специфич-е водор связи, в резте чего осуществл-ся так называемое уотсон–криковское спарив-е. Аденин всегда образ водор связи с тимином, а гуанин с цитозином. Такая законом-ть назыв комплементтью (определ последов-ть оснований в противопол-х цепях ДНК. Данная законом-ть очень важна для репликации ДНК.
3. Самовоспроизведение наследственного материала. Репликация ДНК.
Репликация ДНК-самоудв-е мол-л ДНК,к-ое обычно происх перед дел-ем кл-ки. Во время репликацииматер мол-ла раскручив, и комплемент нити её разъедин(образ репликативн вилка) Формир-е репликат вилки происх под дей-ем ферментов геликазы и топоизомеразы. Геликаза разрыв водор связи между комплемент-ными нуклеотидами и разъедин нити, топоизомераза сним напряж-е, возникающее при этом в мол-ле. Одиночн нити матер мол-лы служат матрицами для синтеза дочерних комплемент-х нитей. С одиночн нитями связыв SSB-белки(дестабилизирующие белки),к-ые не дают им соедин в двойн спираль. В рез-те репликации образ две одинак мол-лы ДНК,полностью повторяющие матер мол-лу. При этом кажд нов мол-ла сост из одной нов и одной стар цепи. Комплемент нити мол-лы ДНК антипараллельны. Наращив-е полинуклеотидной цепи всегда происх в направл от 5' конца к 3' концу. Вследствие этого одна нить лидирующ (3' конец в основании репликативной вилки), а другая - запаздывающая (5' конец в основании вилки) и поэтому строится из фрагменьов Оказаки, растущих от 5' к 3' концу. Фрагменты Оказаки – это участки ДНК, которые у эукариот имеют длину 100-200 нуклеотидов, у прокариот – 1000-2000 нуклеотидов.
Синтез цепи ДНК осуществляет фермент ДНК-полимераза. Она наращив дочерн цепь, присоединяя к её 3' концу нуклеотиды, комплементарные нуклеотидам материнской цепи. Особ-ть ДНК-полимеразы сост в том, что она не может начать работу на «пустом месте», не имея 3' конца дочерней нити. Поэтому синтез лидирующей нити и синтез каждого фрагмента Оказаки начинает фермент праймаза. Это разновидность РНК-полимеразы. Праймаза способна начать синтез новой полинуклеотидной цепи с соедин-я двух нуклеотидов. Праймаза синтезирует из РНК-нуклеотидов короткие затравки - праймеры. Их длина около 10 нуклеотидов. К 3' концу праймера ДНК полимераза начин присоединять ДНК-нуклеотиды.
Фермент экзонуклеаза удал праймеры. ДНК-полимераза достраивает фрагменты Оказаки, фермент лигаза сшивает их.