

Энергетическая функция общего пути катаболизма
.rtfЭнергетическая функция общего пути катаболизма (ОПК)
Конечными продуктами общего пути катаболизма являются СО2, NADН и FADH2. В клетках ОПК является основным поставщиком первичных доноров водорода в ЦПЭ и включает 4 NAD+-зависимые и 1 FAD-зависимую реакции дегидрирования. В этих 5 реакциях, сопряженных с окислительным фосфорилированием АДФ, образуется 14 молекул АТФ (в пересчете на 1 молекулу пирувата). Еще 1 молекулу АТФ поставляет реакция субстратного фосфорилирования АДФ в ходе образования сукцината. Общий выход АТФ составляет 15 молекул АТФ. Распад ацетил-КоА в ЦТК сопровождается синтезом 12 молекул АТФ.
Образованием оксалоацетата завершается один оборот цитратного цикла. В одном обороте цикла лимонной кислоты в 2 реакциях декарбоксилирования происходит образование 2 молекул СО2. В 4 реакциях цитратного цикла происходит дегидрирование с образованием восстановленных коферментов: 3 молекул NADH+H+ и 1 молекулы FADH2. На один оборот цикла затрачивается 2 молекулы воды: одна - на стадии образования цитрата, вторая - на стадии гидратации фумарата.
Восстановленные коферменты (3 молекулы NADH и 1 молекула FADH2), образованные в цикле лимонной кислоты, отдают электроны в ЦПЭ на кислород - конечный акцептор электронов. Восстановленный кислород взаимодействует с протонами с образованием воды.
На каждую молекулу NADH при образовании молекулы воды в процессе тканевого дыхания синтезируются 3 молекулы АТФ, а на каждую молекулу FADH2 - 2 молекулы АТФ.
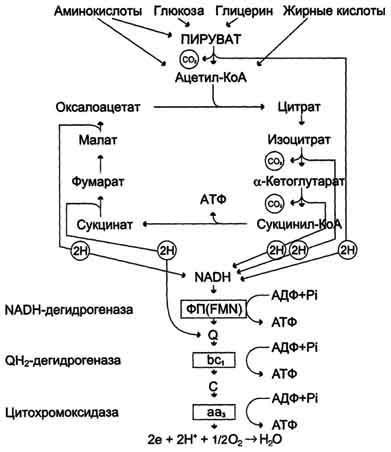
Таким образом, каждый оборот цикла лимонной кислоты сопровождается синтезом 11 молекул АТФ путём окислительного фосфорилирования. Одна молекула АТФ образуется путём субстратного фосфорилирования. В итоге на каждый ацетильный остаток, включённый в цитратный цикл, образуется 12 молекул АТФ.
Регуляция общего пути катаболизма
Скорость ОПК зависит от потребности клеток в АТФ и регулируется с помощью 4 ферментов процесса: пируватдегидрогеназного комплекса (ПДК), цитратсинтазы, изоцитратдегидрогеназы, α-кетоглутаратдегидрогеназного комплекса. Регуляция на уровне ПДК имеет важное значение для обеспечения цитратного цикла "топливными" молекулами ацетил-КоА.
Образование ацетил-КоА из пирувата - необратимый ключевой этап метаболизма. Реакция окислительного декарбоксилирования ПВК в клетке регулируется 2 способами: фосфоролированием/дефосфорилированием и аллостерически.
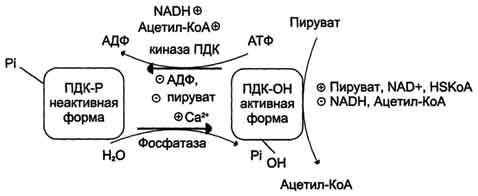
Регуляторным компонентом ПДК является фермент пируватдекарбоксилаза (Е1). Этот фермент имеет аллостерические центры, к которым могут присоединяться активаторы (NAD+, КоА) и ингибиторы (ацетил-КоА, NADH). Кроме трех основных ферментов в состав ПДК входят 2 регуляторных субъединицы - киназа ПДК и фосфатаза ПДК, которые могут соответсвенно фосфорилировать или дефосфорилировать фермент пируватдекарбоксилазу. В дефосфорилированной форме фермент активен и катализирует реакцию окислительного декарбоксилирования пирувата.
Киназа ПДК является аллостерическим ферментом, ее активность зависит от концентрации многих соединений: ацетил-КоА, NADH и АТФ активируют киназу, а NAD+, КоА, пируват, АДФ ее ингибируют. Фосфатаза ПДК активируется Са2+.
В ЦТК цитратсинтаза, изоцитратдегидрогеназа и α-Кетоглутаратдегидрогеназный комплекс являются аллостерическими ферментами. Их активность зависит от соотношения АТФ/АДФ и NADH/NAD+ в клетке. Высокие концентрации АТФ и избыток NADH понижают их активность. В регуляции принимают промежуточные метаболиты ЦТК – субстраты и продукты реакций. Так, цитратсинтаза активируется оксалоацетатом и ингибируется при повышении концентрации цитрата и сукцинил-КоА в митохондриях. Сукцинил-КоА также является ингибитором и α-кетоглутаратдегидрогеназного комплекса.
Самую медленную ренакцию процесса катализирует изоцитратдегидрогеназа, поэтому при повышении в клетке концентрации АТФ и NADH активность этого фермента снижается в наибольшей степени.
В мышцах при физической работе повышается содержание Са2+, который является важным регуляторным фактором скорости ОПК. Са2+ активирует фосфатазу ПДК, которая дефосфорилируется и активирует ПДК. Са2+ активирует изоцитратдегидрогеназу и α-Кетоглутаратдегидрогеназный комплекс.
Сукцинатдегидрогеназа не регулируется аллостерическим путем, но способна ингибироваться при повышении концентрации малоновой, щавелевой и глутаровой кислот.
Анаболические функции цитратного цикла
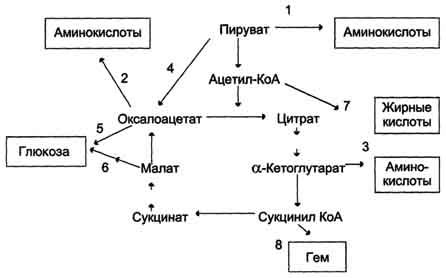
ОПК может выполнять в организме анаболическую функцию, так как многие метаболиты, участвующие в этом процессе, служат субстратами для синтеза различных соединений.
-
из пирувата может синтезироваться аланин
-
из ацетил-КоА = жирные кислоты и холестерол
-
α-кетоглутарат используется для синтеза глутамата, глутамина и пролина
-
сукцинил-КоА является предшественником гема
-
оксалоацетат используетя для синтеза аспартата, аспаратина и глюкозы