

Слижевский Гидробионика в судостроении
.pdfГЛАВА 2 |
31 |
|
|
особенности дельфины, которые помимо высоких скоростных ка- честв обладают рядом удивительных психофизических особенностей, таких, как умение слушать, издавать звуки различного характера, запоминать те или иные явления, а также доверчивым расположением к людям и легкостью, с которой подвергаются дрессировке. Гидродинамические исследования китообразных касались формы тела с точки зрения возможности использования ее особенностей при проектировании подводных лодок. Практическим результатом этих исследований явилась форма корпуса экспериментальной ПЛ "Albacore", спущенной на воду в 1953 году. В дальнейшем аналогич- ная форма нашла применение при проектировании как зарубежных, так и отечественных атомных ПЛ.
Систематические исследования особенностей плавания дельфинов были начаты в 1960 году, когда военным ведомством США было осуществлено финансирование научной программы, целью которой явились дрессировка дельфинов для использования их в каче- стве помощников в освоении океана, а также определение максимальной скорости их плавания. Первые наблюдения за дрессированными дельфинами подтвердили существующее мнение о том, что они способны двигаться с большими скоростями в течение длительного времени. В связи с этим была поставлена задача изучения механизма движения их тела и физиологических особенностей, которые обеспечивают высокоскоростные качества при сравнительно малых затратах энергии.
Уместно отметить, что впервые несоответствие между мускульной мощностью китообразных и достигаемыми скоростями было обнаружено еще в 1963 году известным английским зоологом проф. Д.Греем. Сопоставляя мощность, затрачиваемую на движение твердых тел, подобных телу дельфина, Д.Грей пришел к заключению, что при скорости 20 уз дельфин должен затрачивать мощность, которая почти в 10 раз превышает мощность его мускулатуры. Это несоответствие получило в дальнейшем наименование "парадокса Грея" или "эффекта Грея".
Одним из возможных объяснений этого парадокса явилось предположение, что дельфины, которые движутся со скоростями, соответствующими турбулентному режиму обтекания, обладают способностью ламинаризировать пограничный слой и что эту способность следует искать в особых свойствах кожного покрова, а также в мускульной и сосудистой системах дельфинов.
32 |
ГИДРОБИОНИКА В СУДОСТРОЕНИИ |
|
|
Биоэнергетические затраты животных и в том числе гидробионтов в настоящее время могут быть с достаточной степенью точности оценены по обмену веществ (метаболизму), с которым связаны все жизненные процессы. При этом учитывают реагирующие компоненты и их калорийность, прежде всего потребление кислорода. Рассматривая гидробионта в плавании как живую машину, среднюю мощность общего обмена N можно назвать физиологически располагаемой мощностью, которая химически высвобождается из субстраты организма посредством метаболизма. При этом гидродинамически эффективная мощность Ne = Rν , затрачиваемая на движение с постоянной скоростью υ0 (ãäå R –0 средняя сила сопротивления), связана с мощностью активного обмена Na следующим соотношением:
Ne = ηèäηìNa, |
(2.1) |
ãäå ηèä – идеальный гидродинамический КПД; ηì ≈ 0,2 – КПД мышц, который учитывает эффективность превращения биохимической энергии гидробионта в механическую энергию мышц Nì.
В свою очередь, согласно открытому еще в 1915 году Броуди принципу
N = kN0, |
(2.2) |
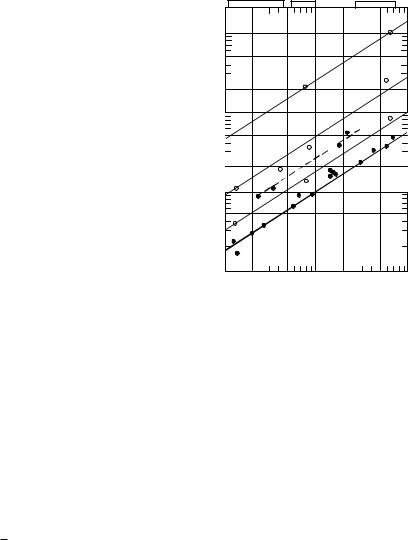
ãäå N0 – средняя мощность основного обмена, затрачиваемая на деятельность гидробионта в состоянии покоя; k – коэффициент кратности, определяемый характером выполняемой животным работы, ее активностью и продолжительностью. При этом активная мощность Na млекопитающего может превышать основной обмен в k = 1,6; 4,8 и 25 раз (рис.2.1), в связи с чем
Na = N – N0 = (k – 1)N0. |
(2.3) |
Как следует из рис.2.1, основной обмен у афалин соответствует таковому у человека (линия 1). Однако в возбужденном состоянии дельфинов (афалины и морской свиньи) их основной обмен превышает норму более чем в два раза (линия 1′).
Другим, не менее эффективным способом оценки биоэнергети- ческих затрат гидробионта, является их определение на основе обработки кинограмм прыжка из воды скоростных водных животных –
ГЛАВА 2 |
33 |
|
|
дельфинов, рыб и кальмаров. |
N0, Na, Âò |
Ñ |
|
× |
|
Ë |
|
Так, например, по результатам |
2 |
|
|
|
|
|
|
обработки кинограмм прыжков |
104 |
|
|
|
|
|
|
была определена максимальная |
|
|
|
|
|
|
|
отдача мощности дельфина L = |
5 |
|
|
|
|
|
|
|
|
|
4 |
|
|
|
|
= 2,0 м и m = 80 кг, которая со- |
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
ставила Ne = 3 кBт при скоро- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
сти выхода из воды υ0 = 9 ì/ñ. |
103 |
|
|
3 |
|
|
|
|
|
|
|
|
|
|
|
|
5 |
|
|
Àô |
|
|
|
|
|
|
|
|
|
||
|
2 |
|
|
1¢ |
|
|
|
|
|
|
|
|
|
|
|
|
2 |
ÌÑ |
|
1 |
Àô |
|
|
|
|
|
|
|
|||
|
10 |
|
|
|
|
|
|
Рис.2.1. Зависимость мощности |
5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
основного и активного обмена |
2 |
|
|
|
|
|
|
от массы бионта |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10 |
2 |
5 |
102 |
2 |
5 |
103 |
|
|
|
|
|
|
|
m, êã |
2.2. Сопротивление воды движению гидробионтов. Физические причины и явления, сопровождающие возникновение сопротивления
Тело гидробионта в общем случае совершает пространственное движение в безграничной вязкой жидкости. Независимо от характера движения за счет взаимодействия поверхности тела гидробионта с окружающей жидкостью в каждой точке этой поверхности возникает гидродинамическая реакция в виде поверхностных сил, напряженность которых
|
n = − |
|
0 p + τ 0τ , |
(2.4) |
p |
n |
– орты внешней нормали и касательной в произвольной
точке А(x,y) поверхности гидробионта; p, τ – нормальное и касательное напряжения поверхностных сил гидродинамической реакции.
При этом главный вектор гидродинамических сил, действующих
34 |
ГИДРОБИОНИКА В СУДОСТРОЕНИИ |
|
|
на поверхность тела гидробионта, будет
R = ∫ pn dS = − ∫ pn 0 p+ ∫ ττdS,
SS
ãäå Rp = − ∫ pn0 dS – главный вектор сил давления; RF
S
главный вектор сил трения.
Проекция Rx = R главного вектора гидродинамических сил на направление продольного движения именуется силой сопротивления продольному движению гидробионта.
Рассмотрим особенности сопротивления движению гидробионтов, которое, как и для твердых тел, в соответствии с парадоксом Даламбера–Эйлера возникает за счет сил вязкости жидкости и нестационарности движения гидробионта.
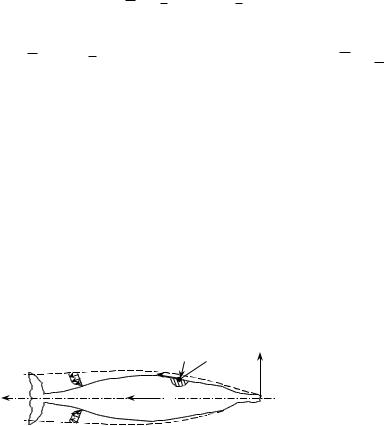
Для выяснения физических причин и явлений, сопровождающих возникновение сопротивления, будем рассматривать равномерное поступательное движение твердого тела, имеющего форму гидробионта (рис.2.2). В этом случае на поверхности тела формируется тонкий пограничный слой, в котором проявляется действие сил вязкости и который является причиной возникновения вязкостного сопротивления движению и сопровождающих его явлений.
t |
p |
A(x,y) |
y |
|
|
|
x |
R |
ds |
dS |


 0
0
Рис.2.2. Распределение характеристик погранич- ного слоя вдоль поверхности тела гидробионта
Во всех точках на поверхности тела относительная скорость обтекания равна нулю (ньютоновская гипотеза прилипания). В результате этого возникают касательные напряжения вязкости τ, проекция главного вектора которых на направление продольного движения равна силе сопротивления трения
RF = ∫ τ cos( τ 0 , x ) dS .
S
Наличие пограничного слоя приводит, кроме того, к перерас-
|
ГЛАВА 2 |
35 |
пределению давления вдоль поверхности тела по сравнению с иде- |
||
альной жидкостью (рис.2.3). |
|
u0 |
|
|
|
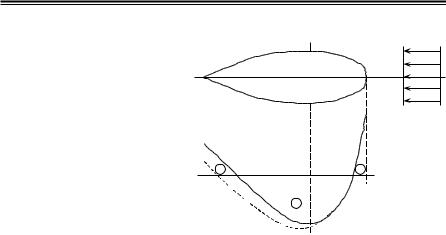
Рис.2.3. Закон распределения |
|
Dp = p-p0 |
избыточного давления ∆p ïî |
|
|
длине тела: |
+ |
+ |
––––––– – идеальная жидкость; |
|
_ |
- - - - - - – вязкая жидкость |
|
|
|
|
|
Так как в идеальной жидкости проекция главного вектора сил давления на направление движения тела равна нулю (парадокс Да- ламбера–Эйлера), то за счет понижения давления в кормовой оконечности, обусловленного влиянием пограничного слоя, возникает сопротивление давления, именуемое сопротивлением формы,
Rp = − ∫ pcos( n0, x )dS .
S
Таким образом, сопротивление движению будет
R = RF + Rp
и может быть вычислено по следующей формуле:
R = C |
ρυ |
20 |
S , |
(2.6) |
|
|
|||
2 |
|
|
|
|
где ρ – плотность воды; υ0 – скорость гидробионта; S – смоченная поверхность.
Входящий в (2.6) коэффициент сопротивления C = C(Re,Φ) для твердого тела, имеющего форму поверхности тела гидробионта и движущегося с постоянной скоростью в безграничной вязкой жидкости, является функцией числа Re = υ0L/ν и формы поверхности тела (Φ – семейство безразмерных геометрических параметров формы тела).
36 |
ГИДРОБИОНИКА В СУДОСТРОЕНИИ |
|
|
Соотношение между составляющими сопротивления RF è Rp зависит главным образом от формы поверхности тела. Так, для скоростных гидробионтов, имеющих удлиненную хорошо обтекаемую форму поверхности тела, главным и определяющим общее сопротивление является сопротивление трения RF. В свою очередь, сопротивление трения зависит от режима течения вязкой жидкости в пограничном слое – ламинарного (послойного) либо турбулентного (бурного течения), сопровождающегося переходом частиц жидкости из слоя в слой.
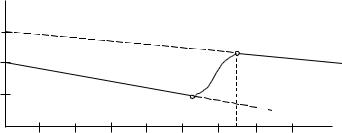
Характерный вид зависимости C(Re,Φ) в логарифмических координатах для хорошо обтекаемых твердых тел приведен на рис.2.4.
lgÑ |
|
|
|
|
|
|
|
|
|
|
Турбулентный |
||
|
Ламинарный |
|
|
|
|
|
1 |
2 |
3 |
4 |
5Reêðí 6 Reâêð7 |
8 |
lgRe |
Рис.2.4. Зависимость коэффициента сопротивления C = C (Re) для |
||||||
|
хорошо обтекаемого твердого тела |
|
|
|||
Анализ диаграммы (рис.2.4) позволяет выделить следующие три режима обтекания твердого тела:
первый при Re ≤ Re íêð ( Re íêð – нижнее критическое значение числа Re, при котором наблюдается переход турбулентного режима обтекания в ламинарный). В этом диапазоне чисел Re наблюдается устойчивый ламинарный режим обтекания, характеризуемый относительно малым по сравнению с турбулентным режимом коэффициентом сопротивления;
второй при Re íêð ≤ Re ≤ Re âêð . В этом диапазоне чисел Re наблюдается неустойчивый режим обтекания, когда в носовой оконечности возникает ламинарный участок, длина которого уменьшается с увеличением числа Re;
третий при Re ≥ Re âêð ( Re âêð – верхнее критическое значение чис-
ГЛАВА 2 |
37 |
|
|
ла Re, при котором наблюдается переход ламинарного режима обтекания в турбулентный). В этом диапазоне по всей поверхности твердого тела наблюдается турбулентный режим обтекания, характеризуемый относительно большим по сравнению с ламинарным режимом коэффициентом сопротивления.
Для скоростных рыб и китообразных числа Рейнольдса составляют Re = 106…108, что для твердых тел соответствует турбулентному режиму обтекания и, соответственно, относительно большим зна- чениям коэффициента сопротивления. Однако биоэнергетические расчеты показывают, что сопротивление гидробионтов значительно ниже и одной из главных причин этого является ламинаризация пограничного слоя у активно плавающих гидробионтов, связанная с увеличением длины ламинарного участка и существенным (в несколько раз) снижением коэффициента сопротивления.
Анализ гидробионических особенностей функционально-морфо- логических свойств и кинематики движения китообразных и скоростных рыб позволяет рассматривать в качестве возможных следующие способы ламинаризации пограничного слоя у гидробионтов:
наличие ламинаризованной формы тела; гашение турбулентной пульсации в пограничном слое за счет
упругодемпфирующих свойств кожного покрова гидробионтов; выделение в пограничный слой гидробионта биологической слизи; нестационарный характер движения гидробионта.
2.3. Ламинаризованная форма тела скоростных китообразных и скоростных рыб
Естественное в процессе эволюции развитие ламинаризованной формы тела гидробионтов и искусственное в процессе создания ламинаризованных профилей крыльев и средств движения в воде связаны с устойчивостью ламинарного пограничного слоя, формируемого на поверхности этих тел при движении в вязкой жидкости. Ламинарный пограничный слой считается устойчивым, если возникающие в нем местные возмущения поля скоростей и давлений с тече- нием времени затухают после прекращения воздействия причин, вызвавших эти возмущения. В противном случае происходит потеря устойчивости ламинарного режима течения и его переход в турбулентный.
38 |
ГИДРОБИОНИКА В СУДОСТРОЕНИИ |
|||
Из теории устойчивости ламинарного режима течения извест- |
||||
но, что, наряду с характером и величиной местных возмущений, оп- |
||||
ределяющим является форма эпюры скоростей в направлении, пер- |
||||
пендикулярном потоку, которая, в свою очередь, зависит от особен- |
||||
|
|
|
ностей геометрии повер- |
|
|
Точка |
|
хности тела в продольном |
|
|
перегиба |
|
направлении. Практичес- |
|
|
y |
|
||
|
u0 |
ки важным результатом |
||
|
|
|||
|
|
|
этой теории является вы- |
|
x |
0 |
|
вод о том, что ламинар- |
|
|
|
|
ные потоки, имеющие |
|
|
|
|
точку перегиба на эпюре |
|
|
|
|
скоростей,неустойчивы. |
|
|
Dp |
|
Существование точек |
|
+ |
+ |
|
перегиба на эпюре скоро- |
|
|
стей непосредственно свя- |
|||
|
_ |
|
||
|
|
зано с продольным гради- |
||
|
|
|
||
|
|
|
ентом давления при об- |
|
Рис.2.5. Картина распределения избыточно- |
текании тела (рис.2.5). |
|||
В ламинарном погра- |
||||
го гидродинамического давления ∆р вдоль |
||||
ничном слое в области |
||||
|
поверхности тела |
|
||
|
|
отрицательного градиен- |
||
|
|
|
||
та скорости d ( ∆p ) < 0 (область падения давления) от носика до наи- |
||||
|
dx |
|
|
|
более широкой части тела профили скоростей не имеют точек пере- |
||||
гиба. Это обусловлено тем, что в этой области происходит "разгон" |
||||
потока и поэтому частицы жидкости, движущиеся в пограничном |
||||
слое, обладают большой кинетической энергией в продольном на- |
||||
правлении и, соответственно, большей устойчивостью по отноше- |
||||
нию к различного рода поперечным возмущениям. В области поло- |
||||
жительного градиента давления d ( ∆p ) > 0 от наиболее широкой ча- |
||||
|
|
dx |
|
|
сти тела до кормовой точки эпюра скоростей становится менее пол- |
||||
ной с возможным образованием точки перегиба (см. рис.2.5), а так- |
||||
же за счет дополнительной потери энергии вблизи поверхности тела, |
||||
вызванной влиянием сил вязкости. Поэтому частицы жидкости об- |
||||
ладают малой кинетической энергией в продольном направлении и |
||||

ГЛАВА 2 !'
соответственнно слабой устойчивостью по отношению к попереч- ным возмущениям.
Отсюда следует, что при обтекании тела положение точки минимума давления (наиболее широкая часть) оказывает определяющее влияние на положение точки перехода ламинарного в турбулентный режим течения в пограничном слое. В грубом приближении можно считать, что точка перехода расположения намного ниже по течению, чем точка минимума давления.
Это служит физической основой для создания ламинаризованной формы тела. У таких тел наиболее широкая часть в зависимости от относительной скорости, характеризуемой числом Re = υL/ν и соответствующей переходному режиму обтекания Re íêð ≤ Re ≤ Re âêð , находится на определенном расстоянии от носика тела. Благодаря этому пограничный слой, формируемый при обтекании тела потоком вязкой жидкости, сохраняется ламинарным на значительно большем протяжении, чем у обычных тел, что приводит к существенно-
му снижению вязкостного со- |
|
|
|
|
40 % |
|
|
противления при смешанном |
|
|
|
|
|
16 % |
|
режиме обтекания. Так, для |
|
|
|
некоторых крыльевых профи- |
|
44 % |
à |
лей это уменьшение достига- |
|
|
|
|
|
18 % |
|
ет двукратной величины. |
|
|
|
Исследования американ- |
|
|
á |
ских и немецких ученых по- |
|
45 % |
|
|
% |
||
зволили сделать вывод, что |
|
|
|
|
|
14 |
|
наиболее близкими к ламина- |
|
|
|
|
|
â |
|
ризованным формам являют- |
|
55 % |
|
|
|
||
ся тела форели, тунца, акулы, |
|
|
% |
дельфина, кита и других, име- |
|
|
18 |
|
|
|
|
ющих ламинарный участок, |
|
70 % |
ã |
длина которого зависит от |
|
% |
|
|
|
||
числа Re (рис.2.6) и определя- |
|
|
11 |
|
|
|
|
ется положением по длине |
|
|
|
|
|
ä |
|
гидробионта максимальной |
Рис.2.6. Сопоставление профилей раз- |
||
толщины. |
личных водных животных по располо- |
||
С увеличением числа Re |
жению сечения максимальной толщины: |
||
относительное расстояние xmax |
а – барракуда; б – акула; в – морская собака |
||
от носика до наиболее широ- |
|
(акула); г – щука; д – акула-аллигатор |
|
40 |
ГИДРОБИОНИКА В СУДОСТРОЕНИИ |
|
|
кой части тела гидробионта увеличивается. При этом происходит трансформация формы носа: для относительно малых чисел нос имеет полную эллиптическую форму, с увеличением Re профиль носа приобретает более заостренную параболическую форму. Так, например, для форели (Re = 1,2 106) с достаточно полной эллиптической
формой носа xmax /L = 0,30, в то время как для тунца (Re = = 3,8 107) с заостренной параболической формой носа xmax /L = 0,50. Дельфи-
íû (Re = 2 107), представляющие наибольший интерес в качестве биологического аналога при создании технических средств движения в воде, имеют заостренную форму носа и xmax /L = 0,40.
Весьма эффективным современным средством изучения картины обтекания движущегося тела является метод визуализации с помощью микроскопических люминесцирующих одноклеточных водорослей. Это явление было известно еще древним грекам, наблюдавших за стаей дельфинов, резвящихся в светящемся море.
В дальнейшем было установлено, что свечение водорослей возникает в вязких потоках, если вязкие напряжения в потоке превышают 0,1 Па. На модели дельфина, плывущего со скоростью 2 м/с, было подтверждено, что в пределах пограничного слоя вязкие напряжения превышают указанное значение, сравниваясь с ним на внешней границе. Вот почему за пределами пограничного слоя водоросли не светятся и при движении дельфина можно заметить четко очерченную полосу пограничного слоя. Эти наблюдения, кроме того, позволили обнаружить, что голова дельфина омывается плавными струями ламинарного течения. Последнее, в свою очередь, подтверждает наличие ламинарного участка при обтекании тела дельфина.
Исследования немецких и американских ученых в области гидробионики позволили создать целую группу хорошо обтекаемых и так называемых ламинаризованных профилей, которые получили принятое в США наименование – профили NACA. Так, например, серия профилей NACA 63 близка к форме тела форели, серия NACA 66 разработана в результате изучения формы тела дельфина, а серия NACA 67 – изучения формы тела тунца. Разнообразие форм профилей в пределах одной серии достигается варьированием максимальной толщины профиля. Так, например, два профиля NACA 66 (NACA 66-018 и NACA 66-033) имеют относительную толщину 18 и 33 % соответственно.