

Эволюционное учение
.pdf" . . . я убежден, что Естественный Отбор
был самым важным, но не исключительным, фактором изменения."
Ч. Дарвин
Часть
Учение
О МИКРОЭВОЛЮЦИИ
¥ |
Г - |
Учение о микроэволюции — центральный раздел современного эволюционного учения, рассматривает механизм эволюционного процесса и включает главы, посвященные различным факторам эволюции, эволюционному материалу и
эволюционному |
явлению, |
наконец, виду, |
|||||
видообразованию и |
|
возникновению |
различных |
||||
адаптаций |
как |
главным |
результатам |
|
|||
эволюционного прогресса на этом уровне. |
|||||||
Строгий анализ в любой области возможен |
|||||||
лишь тогда, когда удается вычленить |
единицы |
||||||
и описать |
элементарные |
структурные |
|||||
и протекающие в них процессы. |
|
|
|||||
Он достигнут применительно к |
благодаря |
||||||
микроэволюционным |
событиям |
||||||
использованию |
идей |
и |
подходов |
генетики. |
|||
Микроэволюция изучает факторы и механизмы внутривидовой дифференциации, завершающиеся видообразованием.
Именно здесь реализуется механизм всего грандиозного эволюционного процесса на Земле.
Вклассическом дарвинизме начальные стадии
видообразования оставались неизученными, они как раз составляют основы современного учения о микроэволюции.
Г л а в а 7
|
Возникновение |
учения |
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
о |
минроэволюции. |
|
|
|
|
|
|
|
|
|
|
|
|
||||||
Популяция — элементарная |
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
единица |
эволюции |
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
На основе классических сравнитель- |
отношении |
эволюционного |
процесса. |
|||||||||||||||||
но-анатомических, |
эмбриологических, |
Микроэволюция — |
эволюционные |
|
из- |
||||||||||||||||
биогеографических, |
|
палеонтологичес- |
менения, которые идут внутри вида и |
||||||||||||||||||
ких и других методов во второй полови- |
приводят |
|
к |
его |
дифференцировке, |
||||||||||||||||
не XIX в. были выяснены многие зако- |
завершаясь |
видообразованием^. |
|
||||||||||||||||||
номерности эволюции применительно к |
В 30-е годы в результате интен- |
||||||||||||||||||||
крупным группам организмов в боль- |
сивного развития популяционной ге- |
||||||||||||||||||||
ших (геологические) интервалах вре- |
нетики возникла объективная возмож- |
||||||||||||||||||||
мени |
(макроэволюция). В то же время |
ность более глубокого познания са- |
|||||||||||||||||||
начальные |
стадии |
|
процесса |
эволю- |
мого |
механизма |
возникновения новых |
||||||||||||||
ции — собственно |
механизм |
эволюци- |
признаков (адаптаций) и в конце кон- |
||||||||||||||||||
онного |
процесса, |
действующий |
при |
цов |
механизма |
возникновения |
видов, |
||||||||||||||
возникновении новых видов, оставались |
чем |
это |
было |
|
возможно |
|
раньше, |
||||||||||||||
малоизученными. В этой главе рассмат- |
лишь на основе наблюдений в природе. |
||||||||||||||||||||
ривается более подробно элементарная |
Крайне |
существенным |
моментом |
при |
|||||||||||||||||
эволюционная |
единица, |
без |
которой |
этом |
оказалась |
возможность |
прямого |
||||||||||||||
никакого |
|
эволюционного |
процесса |
эксперимента при изучении механиз- |
|||||||||||||||||
быть не может,— популяция. Но перед |
мов эволюции: благодаря использова- |
||||||||||||||||||||
этим кратко охарактеризуем весь мик- |
нию дрозофилы и других быстрораз- |
||||||||||||||||||||
роэволюционный уровень исследования |
множающихся видов организмов стало |
||||||||||||||||||||
эволюции. |
|
|
|
|
|
|
|
возможным |
экспериментально |
модели- |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
ровать |
эволюционные |
ситуации |
и |
|||||||
7.1. |
Глазная |
особенность |
изучении |
наблюдать |
протекание |
эволюционного |
|||||||||||||||
процесса как бы через «лупу времени». |
|||||||||||||||||||||
|
|
|
микроюолюции |
|
|
||||||||||||||||
|
|
|
|
|
Оказалось, что за короткое время |
||||||||||||||||
|
Как |
известно, |
строгий |
и |
доста- |
экспериментатор |
|
мог получить |
резуль- |
||||||||||||
|
таты |
изменения |
многих |
десятков |
|||||||||||||||||
точно точный анализ в любой области |
|||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
||||||||||||
возможен лишь тогда, когда удается |
|
|
|
|
|
|
|
|
|
|
|||||||||||
вычленить |
и |
описать |
элементарные |
1 Термин «микроэволюция» был использо- |
|||||||||||||||||
структурные |
единицы |
и |
протекающие |
||||||||||||||||||
ван впервые |
Ю. А. Филипченко в 1927 г., |
||||||||||||||||||||
с их участием явления, В большинстве |
для |
того |
чтобы |
подчеркнуть несводимость, |
|||||||||||||||||
разделов |
биологии |
|
такое выделение |
на взгляд автора, процессов эволюции крупного |
|||||||||||||||||
пока |
не |
произведено. Однако |
благода- |
масштаба к процессам видообразования. В |
|||||||||||||||||
ря |
использованию |
генетических |
идей |
современном |
смысле |
термин «микроэволюция» |
|||||||||||||||
впервые был предложен Ф. Г. Добржанским |
|||||||||||||||||||||
и |
подходов |
оно стало |
возможным в |
(1937) и |
Н. В. Тимофеевым-Ресовским (1938). |
||||||||||||||||
|
|
|
|
|
|
Часть |
3. Учение |
о « и к р |
.ззолюцим |
|
|
|
|
|
|
||||
поколений дрозофил или многих сотен |
Популяция |
— |
это |
минимальная |
|||||||||||||||
поколений |
микроорганизмов. |
А |
за |
самовоспроизводящаяся |
группа |
особей |
|||||||||||||
такое время в изучаемых совокуп- |
одного |
|
вида, |
на |
протяжении |
эволю- |
|||||||||||||
ностях удается наблюдать действи- |
ционно |
|
длительного |
|
времени |
населяю- |
|||||||||||||
тельно значимые эволюционные из- |
щая определенное пространство, об- |
||||||||||||||||||
менения — возникновение |
тех |
|
или |
разующая |
самостоятельную |
генетичес- |
|||||||||||||
иных |
признаков |
и |
свойств |
вплоть |
кую систему и формирующая собствен- |
||||||||||||||
до возникновения репродуктивной изо- |
ную |
экологическую |
|
нишу. |
|
|
|
||||||||||||
ляции |
от |
исходной |
формы. |
|
|
|
Из |
этого |
определения |
вытекает, |
|||||||||
|
Такой |
экспериментальный |
подход |
что популяция — это всегда достаточно |
|||||||||||||||
сразу же выделил область изучения |
многочисленная группа особей, на про- |
||||||||||||||||||
механизмов эволюции из всех других— |
тяжении большого числа поколений в |
||||||||||||||||||
по существу описательных — областей |
высокой |
степени |
изолированная |
от |
|||||||||||||||
изучения эволюции. Благодаря воз- |
других аналогичных |
|
групп |
особей. |
|
||||||||||||||
можности использования эксперимента |
Популяция — самая мелкая, |
эле- |
|||||||||||||||||
оказывается возможным строго про- |
ментарная группа особей, из тех, |
||||||||||||||||||
анализировать действие любого желае- |
которым присуща эволюция. Ни су- |
||||||||||||||||||
мого фактора на возникновение адап- |
ществующие |
немногие |
поколения |
||||||||||||||||
тации, до тонкостей описать сам |
«семья», |
«соседство» |
(«neighborhood»), |
||||||||||||||||
механизм |
возникновения |
адаптаций. |
«панмиктическая |
единица» |
или |
другие |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
выделяемые |
в |
генетических |
работах |
||||||
|
|
7.2. Понятие |
«популяция» |
|
мельчайшие |
группировки |
особей, |
ни |
|||||||||||
|
|
|
тем более отдельные особи не обла- |
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
||||||||||
|
В |
любой области |
биологического |
дают собственной «эволюционной судь- |
|||||||||||||||
исследования приходится в конце кон- |
бой»: поскольку в масштабе эволюции |
||||||||||||||||||
цов |
расчленять подлежащий |
изучению |
их существование эфемерно, они поги- |
||||||||||||||||
материал на какие-то единицы, далее |
бают, не успев эволюционно изменить- |
||||||||||||||||||
разумно не подразделимые в контексте |
ся (если же |
на их основе в будущем |
|||||||||||||||||
данной области исследования. В гене- |
разовьются |
более |
|
крупные |
группы |
||||||||||||||
тике такой |
единицей оказывается |
ген, |
особей, то они будут обладать иными |
||||||||||||||||
в |
систематике — вид, |
при |
изучении |
признаками и свойствами). |
|
|
|
||||||||||||
экосистем — биогеоценоз и |
т. |
д. |
Не- |
Указание |
на |
минимальный |
размер |
||||||||||||
сомненно, такое подразделение от- |
вызвано |
необходимостью разграниче- |
|||||||||||||||||
ражает |
дискретную |
природу |
жизни. |
ния популяций от группы популяции, |
|||||||||||||||
В |
эволюционном |
исследовании |
такой |
обладающей всеми далее перечислен- |
|||||||||||||||
далее |
|
непод разделимой |
единицей |
ными свойствами. Надо иметь в виду, |
|||||||||||||||
оказывается |
популяция. |
Наблюдения в |
что минимальный размер для само- |
||||||||||||||||
природе показывают, что особи любого |
воспроизводства на протяжении дли- |
||||||||||||||||||
вида животных, растений или микро- |
тельного числа поколений подразуме- |
||||||||||||||||||
организмов |
распределены в пределах |
вает |
численность, |
|
достаточную |
для |
|||||||||||||
видового ареала неравномерно и плот- |
выживания популяции при ее резких |
||||||||||||||||||
ность видового населения всегда варьи- |
изменениях |
численности |
(достигаю- |
||||||||||||||||
рует. Эта неравномерность может быть |
щих тысячекратных величин). Конеч- |
||||||||||||||||||
выражена в двух формах: в форме |
но, популяция у позвоночных в период |
||||||||||||||||||
«островного» распределения групп осо- |
минимальной численности может со- |
||||||||||||||||||
бей или |
в |
форме «сгущений». Участки |
ставлять |
в редких |
случаях, наверное, |
||||||||||||||
сотносительно высокой встречаевсего несколько взрослых размножаю-
мостью особей данного вида чередуют- |
щихся особей. Но в среднем это |
|||
ся с участками низкой плотности. |
всегда |
ближе |
к тысячам |
особей, |
Такие «центры плотности» населения |
чем к десяткам. Надо иметь в виду» |
|||
каждого вида и называют, как правило, |
что, говоря о численности популяции, |
|||
популяциями. |
имеется |
в виду |
эффективная |
величина |
|
|
|
|
|
|
Глааа 7. Возникновение учения о микроэволюции |
|
|
|
|
|
|
|
|||||||||||||||||
численности |
(Nе), т. е. число |
|
раз- |
|
7.3, |
Основные характеристики |
|
|||||||||||||||||||||||
множающихся |
|
животных; |
оно |
всегда |
|
|
|
|
|
популяции |
|
|
|
|
|
|||||||||||||||
ниже общего числа особей, составля- |
как |
|
эколого-генетической |
системы |
||||||||||||||||||||||||||
ющих |
популяцию. |
|
|
|
|
|
|
|
|
|
Основные |
экологические |
характе- |
|||||||||||||||||
Из сказанного ясно, что не все |
|
|||||||||||||||||||||||||||||
группы особей могут быть названы |
ристики популяции — величина (по за- |
|||||||||||||||||||||||||||||
популяциями. Когда |
«непопуляциями» |
нимаемому пространству и численности |
||||||||||||||||||||||||||||
оказываются |
явно |
внутрипопуляцион- |
особей), возрастная и половая струк- |
|||||||||||||||||||||||||||
ные |
(семьи, демы |
и |
т. |
д.) |
или |
над- |
тура, |
а также популяционная динамика. |
||||||||||||||||||||||
популяционные |
группы |
(группы |
попу- |
|
Популяционный ареал. Собственное |
|||||||||||||||||||||||||
ляций разного ранга вплоть до видо- |
пространство (ареал) оказывается од- |
|||||||||||||||||||||||||||||
вого), |
принципиальных |
затруднений |
в |
ним из важных критериев популяции. |
||||||||||||||||||||||||||
применении данного выше определения |
Несомненно, как часть вида популя- |
|||||||||||||||||||||||||||||
не возникает. Однако в других случаях |
ция |
|
обладает |
определенным |
ареалом. |
|||||||||||||||||||||||||
такие затруднительные ситуации воз- |
Особи вне этого ареала выходят из |
|||||||||||||||||||||||||||||
можны. |
|
|
|
|
|
|
|
|
|
|
|
|
состава |
популяции. |
Конечно, |
ареал |
||||||||||||||
|
Нельзя ли признать популяцией па- |
популяции может расширяться, но для |
||||||||||||||||||||||||||||
ру размножающихся особей? Ведь че- |
этого популяция должна освоить это |
|||||||||||||||||||||||||||||
рез определенное число поколений по- |
новое |
пространство. |
Это |
произойдет |
||||||||||||||||||||||||||
томство |
этих |
немногих |
единичных |
лишь после включения его в обяза- |
||||||||||||||||||||||||||
особей может достигнуть значитель- |
тельную для каждой популяции эколо- |
|||||||||||||||||||||||||||||
ной численности и при благоприятных |
гическую нишу. Пространство (ареал), |
|||||||||||||||||||||||||||||
условиях от них может возникнуть |
занимаемое |
популяцией, |
|
может |
быть |
|||||||||||||||||||||||||
новая популяция. Использование в этих |
неодинаковым как для разных видов, |
|||||||||||||||||||||||||||||
случаях |
понятия |
«форма |
популяцион- |
так и в пределах одного вида. Напри- |
||||||||||||||||||||||||||
ного ранга» |
(А. В. Яблоков), возможно, |
мер, в большом смешанном лесу сред- |
||||||||||||||||||||||||||||
позволит |
избежать |
|
ненужных |
споров |
ней полосы нашей страны, занимаю- |
|||||||||||||||||||||||||
по |
поводу |
точного |
|
определения |
той |
щем более или менее однородный |
||||||||||||||||||||||||
или |
иной |
группировки |
в |
природе, |
участок |
местности, |
в |
определенном |
||||||||||||||||||||||
когда |
эволюционные |
потенции |
|
ее |
месте может расти небольшая группа |
|||||||||||||||||||||||||
очевидны, |
|
но |
предыстория — длитель- |
деревьев, каких-либо кустарников или |
||||||||||||||||||||||||||
ность |
самостоятельного |
самовоспроиз- |
многолетних |
|
трав, |
отделенная |
|
от |
||||||||||||||||||||||
ведения — не |
|
может быть |
доказана. |
других таких же групп особей этого |
||||||||||||||||||||||||||
Введение |
|
такого |
|
понятия |
отразит |
вида расстоянием, обычно непреодо- |
||||||||||||||||||||||||
объективно |
существующую |
порой |
лимым для пыльцы. Такая группа |
|||||||||||||||||||||||||||
расплывчатость границ между «хоро- |
особей образует |
отдельную популяцию |
||||||||||||||||||||||||||||
шими» популяциями и «непопуляция- |
со |
сравнительно |
небольшим |
ареалом. |
||||||||||||||||||||||||||
ми». |
|
|
|
|
|
|
|
|
|
, |
|
|
|
На |
|
большом |
|
(в |
|
сотни |
гектаров) |
|||||||||
|
В |
заключение отметим, |
что |
данное |
пойменном |
|
лугу |
может |
|
существовать |
||||||||||||||||||||
|
популяция |
какого-нибудь |
одного |
рас- |
||||||||||||||||||||||||||
определение |
популяции |
может |
|
охва- |
||||||||||||||||||||||||||
|
тения. Одна |
популяция |
прыткой |
яще- |
||||||||||||||||||||||||||
тить |
все |
|
группы |
организмов, |
кроме |
|||||||||||||||||||||||||
|
рицы |
(Lacerta |
agilis) |
может |
занимать |
|||||||||||||||||||||||||
вирусов. Для этих форм пока не удает- |
||||||||||||||||||||||||||||||
от |
0,1 |
га |
до |
|
нескольких |
гектаров; |
||||||||||||||||||||||||
ся |
сформулировать |
даже |
|
операцио- |
|
|||||||||||||||||||||||||
|
отдельные |
популяции водяной полевки |
||||||||||||||||||||||||||||
нального, |
|
рабочего |
определения |
поня- |
||||||||||||||||||||||||||
|
(Arvicola |
terrestris) |
— от |
|
одного |
до |
||||||||||||||||||||||||
тия «популяция». После формулировки |
|
|||||||||||||||||||||||||||||
нескольких |
десятков |
гектаров и |
т. |
д. |
||||||||||||||||||||||||||
такого определения |
(что |
будет |
воз- |
|||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
можным после специального популя- |
Величина ареала популяции в зна- |
|||||||||||||||||||||||||||||
ционного |
|
исследования |
|
этих |
форм |
чительной мере зависит от степени |
||||||||||||||||||||||||
в |
природе) |
станет |
ясным, |
есть |
ли |
подвижности |
|
особей |
— |
«радиусов |
||||||||||||||||||||
необходимость учета этой формы жиз- |
индивидуальной или, точнее, репродук- |
|||||||||||||||||||||||||||||
ни в определении понятия «популя- |
тивной |
активности» |
(Н. В. Тимофеев- |
|||||||||||||||||||||||||||
ция». |
|
|
|
|
|
|
|
|
|
|
|
|
|
Ресовский). |
Если |
такой |
|
радиус |
неве- |
|||||||||||
106 |
|
|
|
|
|
|
|
|
|
i 22 Часть 3. Учение о микроэволюции |
|
|
|
|
|
|
|
|
|
|
||||||||||
лик, то величина популяционного ареа- |
ляной |
улитки |
|
(Cepaea |
nemoralis) |
— |
||||||||||||||||||||||||
ла |
обычно |
также невелика |
(табл. 7.1). |
лишь 1000 экз. |
|
|
|
|
|
|
|
|
|
|||||||||||||||||
У растений радиус репродуктивной ак- |
|
Кратковременные |
|
группировки, |
||||||||||||||||||||||||||
тивности |
определяется |
расстоянием, |
состоящие даже из большого числа |
|||||||||||||||||||||||||||
на |
которое |
|
могут |
распространяться |
особей, нельзя еще считать популя- |
|||||||||||||||||||||||||
пыльца, |
|
семена |
или |
вегетативные |
циями. Так, например, саджа |
(Syrrhap- |
||||||||||||||||||||||||
части, способные дать начало новому |
tes |
paradoxus) |
|
в |
отдельные |
годы |
||||||||||||||||||||||||
растению. |
|
|
|
|
|
|
|
|
|
|
|
встречается |
сравнительно |
|
большими |
|||||||||||||||
|
Во многих других случаях трофи- |
группами за тысячи километров от |
||||||||||||||||||||||||||||
ческий ареал не совпадает с репро- |
своего обычного |
центральноазиатского |
||||||||||||||||||||||||||||
дукционным (с эволюционно-генети- |
пустынно-полупустынного ареала. Раз- |
|||||||||||||||||||||||||||||
ческой точки зрения нас прежде всего |
множающиеся особи этого вида встре- |
|||||||||||||||||||||||||||||
интересует |
|
репродукционный |
|
ареал). |
чены даже в северо-западной Европе. |
|||||||||||||||||||||||||
Несмотря |
на |
огромный |
трофический |
Однако через одно-два поколения эти |
||||||||||||||||||||||||||
ареал белого аиста (Ciconia ciconia), |
группы исчезают. Не являются попу- |
|||||||||||||||||||||||||||||
обитающего |
летом |
в |
Европе, |
а |
зимой |
ляциями и группы в несколько десят- |
||||||||||||||||||||||||
в Африке, каждая пара птиц возвра- |
ков |
рыжих |
полевок |
|
(Clethrionomys |
|||||||||||||||||||||||||
щается обычно в район своего старого |
glareolus), более или менее устойчиво |
|||||||||||||||||||||||||||||
гнезда, и популяции аистов хотя и |
существующие на протяжении одного- |
|||||||||||||||||||||||||||||
смешиваются |
на |
местах |
зимовок, |
двух поколений. Население |
подобных |
|||||||||||||||||||||||||
но во время размножения занимают |
группировок |
периодически |
обновляет- |
|||||||||||||||||||||||||||
сравнительно |
небольшую |
территорию. |
ся |
или |
полностью |
сменяется |
осенью |
|||||||||||||||||||||||
|
Численность особей |
в |
популяции. |
за |
счет |
мигрирующих |
особей. |
|
Попу- |
|||||||||||||||||||||
|
ляцией |
в |
настоящем |
|
примере |
будет |
||||||||||||||||||||||||
В |
связи |
с |
размерами |
ареала |
попу- |
|
||||||||||||||||||||||||
все |
население |
рыжих |
полевок |
леса, |
||||||||||||||||||||||||||
ляций может значительно изменяться и |
||||||||||||||||||||||||||||||
включающее, видимо, |
несколько тысяч |
|||||||||||||||||||||||||||||
численность |
|
особей |
в |
|
популяциях. |
|||||||||||||||||||||||||
|
|
зверьков. |
Это |
более |
крупное |
объеди- |
||||||||||||||||||||||||
У |
насекомых |
и мелких |
растений |
от- |
||||||||||||||||||||||||||
нение как самостоятельное эволюцион- |
||||||||||||||||||||||||||||||
крытых |
пространств |
численность |
осо- |
|||||||||||||||||||||||||||
ное |
единство |
существует |
длительное |
|||||||||||||||||||||||||||
бей в отдельных популяциях может |
||||||||||||||||||||||||||||||
время. |
|
|
|
|
|
|
|
, |
|
|
|
|
|
|||||||||||||||||
достигать сотен тысяч и миллионов |
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||
особей. С другой стороны, популяции |
|
С вопросами о численности попу- |
||||||||||||||||||||||||||||
животных и растений могут быть |
ляций |
связана |
|
проблема |
минимальных |
|||||||||||||||||||||||||
сравнительно небльшими по численнос- |
численностей. |
|
Минимальная |
|
числен- |
|||||||||||||||||||||||||
ти. |
|
Численность |
популяции |
|
стрекоз |
ность — это |
такая |
численность, |
ниже |
|||||||||||||||||||||
(Leucorrhinia |
|
albifrons) |
на |
одном |
из |
которой популяция неизбежно исчезает |
||||||||||||||||||||||||
озер |
Подмосковья |
достигала |
порядка |
по разным эколого-генетическим при- |
||||||||||||||||||||||||||
30 тыс. особей, а численность десятков |
чинам. Численность популяции, как и |
|||||||||||||||||||||||||||||
изученных популяций прыткой яще- |
остальные популяционные |
характерис- |
||||||||||||||||||||||||||||
рицы |
(Lacerta agilis) |
— от |
нескольких |
тики, варьирует. В каждом конкретном |
||||||||||||||||||||||||||
сотен до нескольких тысяч особей; |
случае минимальная численность попу- |
|||||||||||||||||||||||||||||
численность |
одной |
из популяций |
зем- |
ляции |
окажется |
специфической |
для |
|||||||||||||||||||||||
|
|
|
Т а б л и ц а |
7.1. Величина |
радиуса репродуктивной |
активности |
некоторых |
|
|
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
животных и |
растений |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
Вид |
|
|
|
|
|
|
|
|
|
|
Радиус |
активности |
|
|
|
|
|
|||||||
Виноградная |
улитка |
(Helix |
pomatia) |
|
|
|
Несколько десятков |
метров |
|
|
|
|
|
|
||||||||||||||||
Сельдь |
(Clupea |
harengus) |
|
|
|
|
|
|
Несколько сот километров |
|
|
|
|
|
|
|
||||||||||||||
Песец |
(Alopex |
lagopus) |
|
|
|
|
|
|
|
|
» |
|
» |
|
|
» |
|
|
|
|
|
|
|
|
||||||
Северный олень (Rangifer tarandus) |
|
|
|
|
Более ста километров |
|
|
|
|
|
|
|
|
|||||||||||||||||
Ондатра (Ondatra zibethicus) |
|
|
|
|
|
|
Несколько |
сот |
метров |
|
|
|
|
|
|
|
|
|||||||||||||
Серый |
кит |
(Eschrichtius |
gibbosus) |
|
|
|
|
Несколько |
тысяч |
километров |
|
|
|
|
|
|||||||||||||||
Дуб |
(пыльца) |
(Querciis |
petraea) |
|
|
|
|
Несколько метров |
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Г r,«ss 7, В >зникнэс«ние учения о |
микроэволюции |
|
|
|
|
107 |
|||||||||
разных видов. Вряд ли возможно |
Впервые на большое эволюцион- |
||||||||||||||||||
эволюционно |
длительное |
существова- |
ное |
значение колебаний |
численности |
||||||||||||||
ние популяции с численностью мень- |
особей в популяциях обратил внима- |
||||||||||||||||||
ше нескольких сотен особей. Об этом |
ние С. С. Четвериков (1905), он |
||||||||||||||||||
стоит напомнить в связи с возни- |
назвал их волнами жизни. Эти волны |
||||||||||||||||||
кающей в наше время проблемой |
играют роль одного из факторов |
||||||||||||||||||
сокращения численности многих видов. |
эволюции |
(см. гл. 9), |
резко |
и |
нена- |
||||||||||||||
По |
разным |
подсчетам |
численность |
правленно |
изменяющего |
генетический |
|||||||||||||
очень |
разреженной |
ныне |
|
популяции |
состав |
популяций. |
|
|
|
|
|
||||||||
амурского |
тигра составляет |
около |
Возрастной состав популяции. По- |
||||||||||||||||
двухсот особей. Несмотря на строгие |
пуляция состоит из разных по возрасту |
||||||||||||||||||
меры охраны, возможна ситуация, при |
и полу |
особей. |
Для |
каждого |
вида, |
||||||||||||||
которой случайное уменьшение числен- |
а иногда и для каждой популяции |
||||||||||||||||||
ности |
(пожар, |
наводнение, |
отстрел |
внутри вида характерны свои соотно- |
|||||||||||||||
и др.) |
сократит |
популяцию |
настолько, |
шения возрастных групп. На эти соот- |
|||||||||||||||
что |
оставшиеся |
единицы, |
|
не |
находя |
ношения влияют общая продолжитель- |
|||||||||||||
с достаточной частотой партнеров для |
ность жизни, время достижения по- |
||||||||||||||||||
размножения, вымрут на |
протяжении |
ловой зрелости, интенсивность размно- |
|||||||||||||||||
немногих поколений. |
В |
аналогичном |
жения — особенности, |
|
вырабатывае- |
||||||||||||||
положении могут оказаться и другие |
мые в процессе эволюции как при- |
||||||||||||||||||
крупные животные или редкие расте- |
способления к определенным условиям. |
||||||||||||||||||
ния |
(например, |
орхидея венерин баш- |
Рассмотрим |
несколько |
примеров. |
||||||||||||||
мачок, |
распространенная |
в |
централь- |
||||||||||||||||
Предельно |
проста возрастная |
структу- |
|||||||||||||||||
ных |
районах страны, |
и др.), |
|
||||||||||||||||
|
ра популяций таких млекопитающих- |
||||||||||||||||||
Динамика популяции. Размеры по- |
|||||||||||||||||||
эфемеров, как землеройки (Sorex). |
|||||||||||||||||||
пуляции |
(пространственные |
и |
по чис- |
Весной на свет появляются один-два |
|||||||||||||||
лу |
особей) |
подвержены |
|
постоянным |
приплода, |
взрослые вскоре |
вымирают, |
||||||||||||
колебаниям. Причины динамики попу- |
и к осени вся популяция |
состоит |
|||||||||||||||||
ляций в пространстве и времени |
лишь |
|
из |
молодых |
неполовозрелых |
||||||||||||||
чрезвычайно многообразны и в общей |
животных. К весне все перезимовав- |
||||||||||||||||||
форме сводятся к влиянию биоти- |
шие достигают половой зрелости, и |
||||||||||||||||||
ческих и абиотических факторов. Так, |
цикл повторяется. Значительно слож- |
||||||||||||||||||
на |
одном |
из |
небольших |
островков |
нее |
возрастная |
структура |
популяции |
|||||||||||
уюго-западного побережья Англии стадных крупных млекопитающих. В
обитает в диком виде популяция кро- |
стаде |
белух (Delphinapterus |
leucas) |
||||||
ликов |
(Oryctolagus cuniculus). В период |
есть первая возрастная группа, состоя- |
|||||||
максимальной |
численности |
(осенью |
щая из детенышей этого года рожде- |
||||||
благоприятного по кормовым условиям |
ния; вторая( возрастная группа, вклю- |
||||||||
года) |
общее число кроликов достига- |
чающая подросших детенышей про- |
|||||||
ло 10 ООО, а однажды, после холод- |
шлого года/рождения, третья возраст- |
||||||||
ной и малокормной зимы,— сократи- |
ная группа — половозрелые животные |
||||||||
лось до 100 особей. Еще более |
в возрасте 2—3 лет, но обычно не |
||||||||
значительны сезонные колебания чис- |
размножающиеся и, наконец, четвертая |
||||||||
ленности |
некоторых беспозвоночных |
возрастная группа — взрослые размно- |
|||||||
(мухи, комары и др.); сезонная чис- |
жающиеся особи |
в |
возрасте |
от 4—5 |
|||||
ленность их может колебаться в сотни |
до 16—20 лет. Как и обычно у |
||||||||
тысяч |
и |
миллионы раз! |
|
млекопитающих, |
у |
белух возможно |
|||
Кроме сезонных колебаний числен- |
скрещивание между взрослыми живот- |
||||||||
ными |
нескольких |
поколений. |
|
||||||
ности |
известны |
несезонные |
(периоди- |
|
|||||
ческие |
и |
непериодические) |
изменения |
В еще более яркой форме слож- |
|||||
численности особей в отдельных по- |
ный возрастной состав популяции ха- |
||||||||
пуляциях |
(рис. |
7.1). |
|
рактерен для некоторых древесных рас- |
|||||
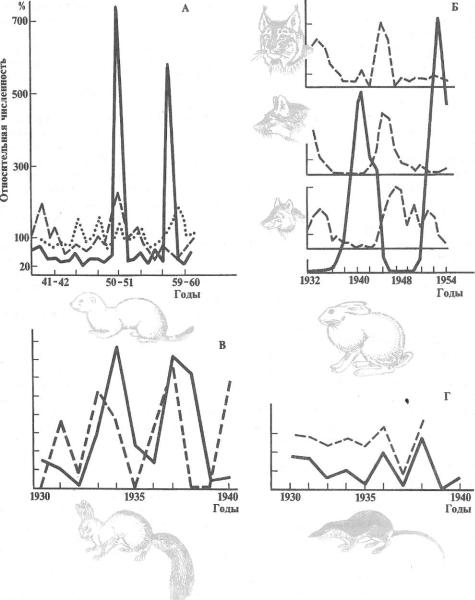
108 i 22 Часть 3. Учение о микроэволюции
Рис. 7.1. Характер колебаний численности особей в |
популяциях некоторых |
млекопитающих: |
||||||||||
А—горностая (Mustela erminea) в трех |
популяциях |
Якутии; |
Б—рыси |
(Felis |
lynx); |
лисицы |
||||||
(Vulpes |
vulpes); |
волка |
(Canis |
lupus) |
и |
зайца |
беляка |
(Lepus |
timidus) |
в |
группе |
|
популяций Севера |
европейской |
части |
СССР; |
В—белки (Sciurus vulgaris) |
в |
Костромской |
||||||
области |
(пунктиром— |
урожай |
семян |
ели); |
Г—обыкновенной |
землеройки |
(Sorex |
araneus) |
||||
(пунктиром—высота весенних паводков) |
(по данным разных авторов из Н. В. Тимофеева- |
|||||||||||
|
|
|
|
Ресовского и др., |
1973) |
|
|
|
|
|
||
тений. |
В |
дубовом |
лесу |
одновремен- |
с мужскими цветками. Личинка червя |
|||||||||||||||
но плодоносят и опыляют друг друга |
Bonellia |
|
viridis |
развивается в |
самку, |
|||||||||||||||
дубы в возрасте более ста лет и совсем |
если после некоторого периода жизни |
|||||||||||||||||||
молодые деревья, которые только- |
в море |
|
она |
не |
сможет |
прикрепиться |
||||||||||||||
только |
созрели. |
|
|
|
|
к другой взрослой самке, если же ли- |
||||||||||||||
|
Половой состав популяции. Извест- |
чинка прикрепится к взрослой самке, |
||||||||||||||||||
но, что генетический механизм опре- |
то из нее всегда получается самец; |
|||||||||||||||||||
деления пола обеспечивает расщепле- |
самцы |
фактически |
паразитируют на |
|||||||||||||||||
ние потомства по полу в отношении |
самках, |
|
они |
в |
десятки |
|
раз |
меньше |
||||||||||||
1:1 |
(первичное |
соотношение |
полов). |
их по размерам и выполняют единст- |
||||||||||||||||
В силу неодинаковой жизнеспособнос- |
венную |
|
|
функцию — оплодотворение |
||||||||||||||||
ти мужского и женского организма |
яиц. |
|
|
|
|
|
|
|
|
|
|
|
||||||||
(разная |
жизнеспособность, несомнен- |
Знание |
экологической |
структуры |
||||||||||||||||
но, эволюционно выработанный при- |
популяции (включающей величину по- |
|||||||||||||||||||
знак) |
это |
первичное |
соотношение |
пуляции и ее динамику в пространстве |
||||||||||||||||
порой уже заметно отличается от |
и во времени, возрастной и половой |
|||||||||||||||||||
вторичного |
(характерно |
при родах у |
состав особей и другие особенности, |
|||||||||||||||||
млекопитающих) и тем более заметно |
изучаемые |
|
популяционной |
эколо- |
||||||||||||||||
отличается от третичного,— характер- |
гией) |
— обязательное |
условие |
глубо- |
||||||||||||||||
ного для взрослых особей. У человека, |
кого изучения популяций как единиц |
|||||||||||||||||||
например, вторичное соотношение по- |
эволюции |
в природе. |
|
|
|
|
|
|||||||||||||
лов составляет 100 девочек на 106 маль- |
Основные |
|
морфофизиологические |
|||||||||||||||||
чиков, к 16—18 годам это соотноше- |
характеристики |
популяции. Поскольку |
||||||||||||||||||
ние (в результате повышенной мужс- |
внутри |
вида |
популяции |
обмениваются |
||||||||||||||||
кой |
смертности) |
выравнивается, и к |
особями и соответственно генетичес- |
|||||||||||||||||
50 годам составляет 85 мужчин на 100 |
ким материалом, то популяции раз- |
|||||||||||||||||||
женщин, а |
к |
80 |
годам — 50 |
мужчин |
личаются |
|
количественным |
соотноше- |
||||||||||||
на |
100 женщин. |
|
|
|
|
нием разных аллелей и частотами |
||||||||||||||
|
И у животных, и у растений |
встречаемости того или иного фено- |
||||||||||||||||||
вторичное |
и |
третичное |
соотношение |
типа, т. е. статистически. Однако не |
||||||||||||||||
полов может колебаться в значитель- |
всегда удается сразу найти !акой |
|||||||||||||||||||
ных пределах у разных видов. У |
признак, |
присутствие |
или |
отсутствие |
||||||||||||||||
некоторых |
|
насекомых |
встречаются |
которого |
|
в |
заметной |
|
концентрации |
|||||||||||
популяции, |
состоящие практически из |
характеризует ту или иную популяцию |
||||||||||||||||||
одних самок в результате сложных |
по сравнению с соседними. В этих |
|||||||||||||||||||
генетических |
процессов, |
влияющих на |
целях проще и надежнее сравнивать |
|||||||||||||||||
элиминацию самцов. Только из самок |
популяции по многим признакам. Осо- |
|||||||||||||||||||
состоят популяции ряда партеногене- |
бенно успешным такой подход бывает |
|||||||||||||||||||
тических видов насекомых и ряда |
при использовании дискретных, качест- |
|||||||||||||||||||
других животных. Интересно заметить, |
венных признаков, которые иногда на- |
|||||||||||||||||||
что |
доля |
партеногенетических самок |
зывают фенами или признаками-мар- |
|||||||||||||||||
в разных популяциях может значи- |
керами генотипического состава попу- |
|||||||||||||||||||
тельно |
варьировать. |
Другой |
крайний |
ляции. Примерами фенов могут слу- |
||||||||||||||||
случай |
в |
соатношении |
полов |
связан |
жить |
красная |
и |
черная |
окраска элитр |
|||||||||||
с эволюционно возникшей исключи- |
у двухточечной божьей коровки, на- |
|||||||||||||||||||
тельно лабильной системой определе- |
личие или отсутствие пленки на зерне, |
|||||||||||||||||||
ния пола. Так, у корнеплода Arisaema |
зазубренная или гладкая ость у пше- |
|||||||||||||||||||
japonica решающий фактор во вто- |
ницы, наличие и расположение отдель- |
|||||||||||||||||||
ричном определении пола — это масса |
ных мелких отверстий в черепе мле- |
|||||||||||||||||||
клубней: самые крупные и хорошо |
копитающих, |
форма |
отдельных |
костей |
||||||||||||||||
развитые клубни дают растения с жен- |
черепа и швов между костями и т. д. |
|||||||||||||||||||
скими цветками, а мелкие и слабые — |
Все |
подобные |
признаки |
отличаются |
||||||||||||||||

110 |
i 22 Часть 3. Учение о м и к р о э в о л ю ц и и |
Рис. 7.2. Пример характеристики популяции по одному фену: места обнаружения устойчивых к антикоагулянту варфарину серых крыс (Rattus
norvegicus) в |
Англии в |
разные годы (по |
Д. |
Друммонду, |
1970) |
качественностью, альтернативностью. Их легко учитывать в любой изучаемой совокупности особей в частотах встречаемости.
В удачных случаях возможно характеризовать популяцию по отдельным, хорошо учитываемым признакам. На рис. 7.2 приведена карта встре-
чаемости в Англии |
серых крыс |
(Rattus norvegicus), не |
чувствительных |
к сильному антикоагулянту варфарину, используемому для борьбы с мелкими грызунами. Генетическая природа этого признака сложна, так что нельзя сказать, что устойчивость к варфарину обусловлена действием какого-то определенного гена. Легкость обнаружения сделала этот физиологический признак удобным признаком-маркёром для определения границ, занимавшихся определенными популяциями до того периода, когда под влиянием бессознательного отбора нечувствительные к
варфарину крысы распространились по всей Англии.
Сопоставление аналогичных данных, полученных по другим признакам, позволяет с большей степенью надежности характеризовать разные популяции, оконтуривать их границы, выяснять степень сходства с соседними популяциями, строить обоснованные гипотезы о путях исторического развития отдельных популяций и их групп, выяснять действие различных эволюционных факторов. В этих случаях приходится морфологические и физиологические признаки описывать, как и при изучении генетики популяции в частотах встречаемости, т. е. подходить к изучению популяций с генетических позиций.
Описанный новый и быстро распространяющийся в наше время подход к изучению морфофизиологических особенностей популяции (фенетический подход) не исключает традиционных способов сравнения популяций по размерам, массе, пропорциям тела составляющих их особей. Следует лишь подчеркнуть, что получаемые традиционными методами результаты крайне трудно использовать для точного эволюционногенетического сравнения популяций изза значительной полигенности и крайне малой наследуемости практически всех размерно-весовых характеристик особей.
Генетическая гетерогенность популяции. С. С. Четвериков (1926), исходя из формулы Харди, рассмотрел реальную ситуацию, складывающуюся в природе. Мутации обычно возникают и сохраняются в рецессивном состоянии и не нарушают общего облика популяции; популяция насыщена мутациями «как губка».
Генетическая гетерогенность природных популяций, как показали многочисленные эксперименты,— главнейшая их особенность. Она поддерживается за счет мутаций, процесса рекомбинации (только у форм с бесполым размножением вся на-