

biohimiyaverstka
.pdf2.1.3. Взаимосвязь обменных процессов
склеточными структурами
Схимической точки зрения обмен веществ – это совокупность большого числа химических реакций, любая из которых может быть воспроизведена вне организма. В отдельности в каждой из этих реакций нет ничего специфически жизненного. Для жизнедеятельности чрезвычайно важно то, что в организме эти отдельные реакции определенным образом сочетаются между собой во времени и в пространстве. Оптимальное течение обмена веществ определяется строгой упорядоченностью химических процессов, которая зависит, с одной стороны, от соотношения скоростей регулируемых ферментами химических реакций, с другой – от степени структурной организации всех элементов организма. Клеточное строение живых организмов – ярчайший пример приспособления структурной организации для выполнения биологической функции.
Каждая клетка представляет собой структурную и функциональную единицу живого организма. Тело некоторых мельчайших организмов состоит из одной клетки (например, одноклеточные бактерии). Другие организмы состоят из множества клеток (в теле человека их несколько сот триллионов), разные типы которых выполняют своим специализированные функции. Например, мышечные клетки способны к сокращению, клетки эпителия выполняют защитную функцию, клетки желез внутренней секреции осуществляют синтез гормона. Однако, несмотря на различия
вспециализации, размерах, форме, в строении клеток разных органов и тканей, а также в способности выполнять различные виды работ имеется много общего. Так, клетки способны запасать и перерабатывать энергию, синтезировать структурные и биологически активные молекулы (белки, ферменты), размножаться, осуществлять различные механические движения, приспособительные реакции, обеспечивающие сохранение основных внутриклеточных параметров при изменении состояния внешней среды.
111
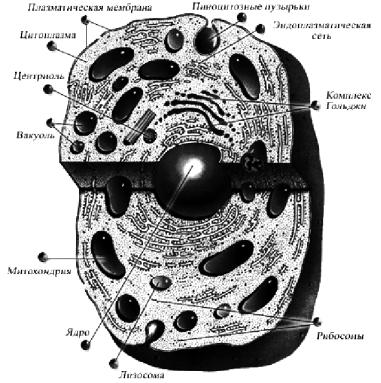
Снаружи клетка окружена оболочкой – трехслойной плазматической мембраной (рис. 28), образованной бимолекулярным слоем липидов, который с обеих сторон покрыт молекулами белка. Экспериментальные данные позволяют предположить, что в белковых слоях мембраны имеются отверстия, «закупоренные» молекулами липидов, расположенными непосредственно под слоем белка. В пользу такой точки зрения свидетельствуют данные о зависимости проницаемости оболочки для разных веществ от размеров их молекул и способности растворяться в липидах.
Рис. 28. Строение клетки
112
Клеточная оболочка обладает избирательной проницаемостью для некоторых веществ – односторонней проницаемостью. Эта особенность обусловлена: наличием особых активных участков в клетке и, кроме того, механизмом, способным совершать осмотическую работу (обеспечивать различия в концентрации ионов по обе стороны мембраны) и потребляющим для этого энергию. Проницаемость клеточной оболочки имеет очень важное значение для протекающих в клетке обменных процессов. Действие многих лекарственных и ядовитых веществ, гормонов заключается в их влиянии на проницаемость клеточных мембран.
Клеточная мембрана образует ряд выпячиваний, которые уходят в глубь клетки. Эти каналы связаны со сложной сетью пузырьков и клеточными органеллами. Образующаяся сложная система взаимосвязанных каналов, пузырьков, полостей получила название эндоплазматической сети или эндоплазматического ретикулума. Эту сеть можно считать транспортной системой клетки, обеспечивающей перенос веществ из окружающей среды и обратно и сообщение между внутриклеточными органеллами.
Снаружи вдоль стенок эндоплазматической сети располагается множество плотных шарообразных зернышек, называемых рибосомами. Рибосомы являются местом синтеза большинства белков клетки. Содержание рибосом теснейшим образом связано с интенсивностью обменных процессов в клетке. Наиболее высоко содержание рибосом в клетках, вырабатывающих белковую продукцию.
В центральной части клетки (исключение составляют мышечные клетки, клетки жировой ткани) располагается ядро. Оно является местом хранения и репликации большей части наследственного материала клетки, который концентрируется в хромосомах, состоящих из ДНК, связанных с белками и небольшим количеством РНК. Кроме хромосом в ядре находится одно или несколько ядрышек – образований, богатых РНК.
113
Ядро отделено от цитоплазмы оболочкой, имеющей поры, через которые осуществляются контакты ядра с цитоплазмой.
Внутриклеточные органеллы – митохондрии – имеют удлиненную форму, диаметр от 0,5 до 1 мкм, длину от 1 до 7–10 мкм. Это сравнительно крупные образования, они видны в обычный микроскоп. Их биологическая функция стала очевидной сравнительно недавно, когда научились выделять их в чистом виде. Митохондрии представляют собой «энергетические станции» клетки, в которых продукты превращений углеводов, жиров, белков окисляются до конечных продуктов обмена (СО2 и Н2О), а освобождающаяся при этом энергия используется для ресинтеза АТФ, доставляющей затем эту энергию во все участки клетки. Количество митохондрий в клетке колеблется в довольно широких пределах (от 50 до 5000) и тесно связано с ее функциональной активностью и энергетическими потребностями.
Отличительной особенностью митохондрий является то, что они имеют самостоятельную генетическую систему
(содержат все известные типы нуклеиновых кислот, составляющих около 1 % сухого веса частиц) и аппарат для синтеза белка (в том числе рибосомы), что делает их в известной мере независимыми от остальной части клетки и обеспечивает им способность к самоудвоению. Благодаря этому содержание митохондрий в мышечных клетках может заметно увеличиваться под влиянием спортивной тренировки. Аналогичные изменения происходят в клетках других органов и тканей, несущих большую функциональную нагрузку при физической работе.
Кроме митохондрий в клетке имеются лизосомы – сравнительно небольшие образования, содержащие гидролитические ферменты, способные расщеплять белки, липиды, углеводы. Функции лизосом особенно ярко проявляются после гибели клетки: гидролитические ферменты высвобождаются из лизосом, и вслед за этим клетка очень быстро растворяется. В клетке лизосомы осу-
114
ществляют «переваривание» фрагментов разрушающихся клеточных структур. Особенно хорошо развиты лизосомы в лейкоцитах, основная функция которых – переваривание попавших в организм извне инородных частиц или организмов (например, бактерий), или веществ самого организма.
Установлено, что вредное действие на организм токсинов, излучений, гипоксии, приводящее к изменениям в тканях, осуществляется через лизосомальный аппарат клетки.
Пространство, ограниченное клеточной мембраной, заполнено цитоплазматическим матриксом, или цитоплазмой. Это водно-дисперсная система, состоящая главным образом из белковых макромолекул в разных стадиях агрегации. Она служит своего рода поддерживающей средой для клеточных органелл. Обычно цитоплазма вблизи наружной клеточной мембраны более плотная и по своим свойствам ближе к твердому веществу, чем около внутреннего слоя.
Кроме белковых молекул в цитоплазме находятся аминокислоты, различные ионы, энергетические субстраты (АТФ, креатин-фосфат, жировые капли, глыбки гликогена) и другие вещества.
2.1.4.Регуляция процессов обмена веществ в организме
Живой организм представляет собой сложнейшую систему, состоящую из огромного количества клеток (организм человека содержит несколько сот триллионов клеток), являющихся, в свою очередь, сложнейшими образованиями. В каждую секунду в организме происходят сотни разнообразных химических реакций, самостоятельных
иобъединенных в процессы. Эти химические реакции и процессы составляют основу жизнедеятельности живого организма. И вся эта сложнейшая система работает четко
ислаженно, быстро и точно, в соответствии с потребнос-
115
тями реагирует изменениями обмена веществ на внешние воздействия, сдвиги во внутренней среде.
Наряду с механизмами активной регуляции обменных процессов, организм характеризуется рядом структурных особенностей и других свойств, способствующих упорядоченности обменных процессов, облегчающих осуществление регуляторных воздействий. К их числу можно отнести прежде всего систему дифференцировки клеток. Клетки одного органа, ткани имеют одинаковое строение, один и тот же набор хромосом.
«Обратный» процесс не обязательно является точной копией «прямого» пути, отдельные его этапы могут обеспечиваться совсем другими реакциями.
К факторам, способствующим эффективной регуляции обмена веществ, можно отнести объединение отдельных химических реакций в процессы, что достигается соответствующей локализацией ферментов.
Ферменты, катализирующие последовательные реакции процесса, располагаются в непосредственной близости, так что продукт одной ферментативной реакции сразу становится субстратом другой.
Регуляция обмена веществ в организме осуществляется нервной и гормональной системами, а также системой автоматической регуляции (саморегуляция).
Основной принцип регуляции обмена веществ в организме сводится к избирательному, точно соответствующему потребностям организма изменению скорости отдельных химических реакций, процессов, комплексов процессов. Хотя этот принцип реализуется в организме достаточно многообразно, в конечном итоге различные механизмы регуляции в подавляющем своем большинстве оказывают влияние на скорость химических реакций путем воздействия на соответствующие ферменты.
Нервная и гормональная системы, системы саморегуляции свое регулирующее влияние на скорость химических реакций осуществляют преимущественно через:
116
1)доступность субстратов (концентрация реагирующих веществ);
2)изменение активности ферментов;
3)изменение количества ферментов;
4)доступность кофакторов.
Одни из этих путей регулирующих воздействий (доступность субстратов и кофакторов, изменение активности ферментов) можно отнести к механизмам срочной регуляции, способным изменить скорость обменных процессов в организме в считанные секунды или минуты. Другие (изменение количества ферментов) осуществляют свое регулирующее воздействие сравнительно медленно. Для проявления их действия требуется несколько часов или даже дней.
Доступность субстрата как регулирующий фактор.
Концентрация субстратов влияет на скорость реакции непосредственно или путем воздействия на активность ферментов. Уменьшение концентрации субстрата приводит к снижению скорости реакции. Когда фермент еще насыщен субстратом, это влияние проявляется исключительно через воздействие на активность ферментов. По мере снижения концентрации субстрата наступает фаза, когда фермент перестает насыщаться субстратом. С этого момента преимущественное влияние на скорость реакции оказывает непосредственно концентрация субстрата.
Аналогичным образом, но в противоположном направлении влияет на скорость химической реакции повышение концентрации субстрата.
Регуляция концентрации субстрата может осуществляться на этапе поступления его в клетку. В качестве регулятора проницаемости клеточной оболочки нередко выступают гормоны. Примером может служить один из путей воздействия гормона инсулина на скорость синтеза гликогена в печени. Инсулин, в частности, повышает проницаемость оболочек клеток печени для глюкозы, увеличивая тем самым скорость поступления глюкозы в клетки, и создает предпосылки для более энергичного синтеза гликогена.
117
В нормальных условиях жизнедеятельности организма большинство ферментов, участвующих в промежуточных реакциях метаболизма, не проявляет своей максимальной активности из-за отсутствия необходимого количества субстратов реакций. Учитывая это, в практике спорта применяются воздействия на скорость аэробного окисления в период восстановления дополнительным введением промежуточных продуктов цикла трикарбоновых кислот: лимонной, янтарной, яблочной.
Следует, однако, заметить, что относительное постоянство состава внутренней среды всего организма и отдельных клеток в отношении субстратов метаболических путей дает основания предполагать, что регуляция за счет изменения доступности субстрата не может изменять скорость химических реакций в широком диапазоне. По-ви- димому, этот механизм как основной не слишком распространен у высших животных. Однако при спортивной деятельности он может играть достаточно серьезную роль. Снижение содержания энергетических субстратов (креатин-фосфата, гликогена) при работе может явиться одной из главных причин замедления скорости ресинтеза АТФ и в конечном итоге падения работоспособности. Непосредственной причиной снижения интенсивности и даже прекращения работы является понижение концентрации АТФ в мышечных волокнах, в клетках центральной нервной системы. АТФ является непосредственным энергетическим субстратом многих энергоемких химических реакций, обеспечивающих формирование двигательного импульса, работу кальциевого насоса, взаимодействие актиновых и миозиновых нитей, приводящее к укорочению мышечного волокна. Концентрация АТФ при ее понижении ниже критического для клетки уровня выступает как главный фактор регуляции скорости этих реакций.
Регуляция скорости химических реакций путем воздействия на активность ферментов является одним из самых главных механизмов. Если с помощью такого механизма регулируется скорость химического процесса,
118
то воздействию подвергается только один фермент, так называемый регуляторный. Как правило, это один из ферментов, катализирующий начальные реакции процесса. В случае разветвления процесса регуляторным является фермент, катализирующих первую реакцию после разветвления метаболического пути. Такой принцип регуляции предотвращает накопление промежуточных продуктов.
Активность регуляторного фермента значительно ниже активности других ферментов, обладающих своеобразной избыточной активностью. Поэтому изменение скорости регулируемой таким ферментом реакции определяет скорость всего процесса в целом.
Данный механизм регуляции является одним из наиболее быстрых и обеспечивает изменение скорости химической реакции в широком диапазоне. Он характеризуется высокой точностью ответной реакции.
Существует много факторов, могущих воздействовать на активность регуляторных ферментов. Воздействие одного из них – концентрация субстрата – уже рассматривалось в предыдущем разделе. Другим фактором может являться концентрация продукта реакции (процесса). Высокие концентрации продукта нередко оказывают ингибирующее влияние на регуляторный фермент. Это воздействие может сказываться по механизму обратной связи, т. е. путем непосредственного воздействия продукта на фермент, или путем изменения рН внутренней среды. Так, в частности, влияет на скорость гликолиза продукт этого процесса – молочная кислота. И в том и в другом случае достигается одно и то же: предотвращение накопления продукта, предотвращение резких изменений во внутренней среде.
Имеются примеры, когда продукт реакции оказывает активирующее влияние на регуляторный фермент или сам выступает в роли катализатора, ускоряя свое образование. Так, один из предшественников ферментов белкового пищеварения – трипсиноген – превращается в свою
119
активную форму – трипсин – под действием энтерокиназы. Энтерокиназа обладает низкой активностью и сравнительно медленно осуществляет превращение трипсиногена в трипсин. Однако образующийся в ходе этой реакции трипсин выступает не только в качестве фермента белкового пищеварения, но и оказывает влияние на трипсиноген, быстро завершая его превращение в трипсин.
Такой механизм регуляции получил название «автокатализ». Он используется там, где нужно быстро осуществить полное превращение субстрата в продукт.
Воздействие на активность ферментов является одним из главных механизмов регулирующего влияния гормонов на обмен веществ. Так, например, адреналин стимулирует расщепление гликогена в печени, мобилизацию жира в жировых депо путем воздействия на активность соответствующих ферментов.
Регуляция обмена веществ путем изменения количества ферментов. Это сравнительно медленный механизм регуляции, для его проявления требуются часы или даже дни. Однако он характеризуется большими возможностями ответной реакции. Он позволяет организму изменять относительные количества и типы ферментов, действующих на любом участке метаболических путей в зависимости от сигналов из окружающей среды. Количество любого фермента может регулироваться на этапе его синтеза или распада. Регуляция на уровне синтеза имеет наибольшее значение.
Вещества, повышающие скорость синтеза ферментов и тем самым увеличивающие их количество в клетке, называются индукторами, угнетающие синтез – репрессорами.
Индукция и репрессия синтеза ферментов осуществляется на уровне генетического аппарата клетки (ДНК или РНК) и заключается в активации или репрессии соответствующих генов, а также синтетической активности рибосом. Регуляция на уровне генетического аппарата может привести:
120