

2 курс / Нормальная физиология / potentsial_deystvia
.docxМембранный потенциал действия. Если на нерв или мышцу нанести раздражение выше порога возбуждения, то МПП нерва или мышцы быстро уменьшится и на короткий промежуток времени (миллисекунда) произойдет перезарядка мембраны: ее внутренняя сторона станет заряженной положительно относительно наружной. Это кратковременное изменение МПП, происходящее при возбуждении клетки, которое на экране осциллографа имеет форму одиночного пика, называется мембранным потенциалом действия (МПД).
МПД в нервной и мышечной тканях возникает при снижении абсолютной величины МПП (деполяризации мембраны) до некоторого критического значения, называемого порогом генерации МПД. В гигантских нервных волокнах кальмара МПД равен - 60 мВ. При деполяризации мембраны до -45 мВ (порог генерации МПД) возникает МПД (рис. 1.15).

действия нервного волокна (А) и изменение проводимости мембраны для ионов натрия и калия (Б).
50
Во время возникновения МПД в аксоне кальмара сопротивление мембраны уменьшается в 25 раз, с 1000 до 40 Ом.см2, тогда как электрическая емкость не изменяется. Указанное снижение сопротивления мембраны обусловлено увеличением ионной проницаемости мембраны при возбуждении.
По своей амплитуде (100-120 мВ) МПД на 20-50 мВ превышает величину МПП. Другими словами, внутренняя сторона мембраны на короткое время становится заряженной положительно по отношению к наружной, — "овершут" или реверсия заряда.
Из уравнения Гольдмана следует, что лишь увеличение проницаемости мембраны для ионов натрия может привести к таким изменениям мебранного потенциала. Значение Ек всегда меньше, чем величина МПП, поэтому повышение проницаемости мембраны для К+ будет увеличивать абсолютное значение МПП. Натриевый равновесный потенциал имеет знак "плюс", поэтому резкое увеличение проницаемости мембраны для этих катионов приводит к перезарядке мембраны.
Во время МПД увеличивается проницаемость мембраны для ионов натрия. Расчеты показали, что если в состоянии покоя соотношение констант проницаемости мембраны для К+, Na+ и СГ равно 1 :0,04:0,45, то при МПД - Рк : PNa : Р = 1 : 20 : 0,45. Следовательно, в состоянии возбуждения мембрана нервного волокна не просто утрачивает свою избирательную ионную проницаемость, а, напротив, из избирательно проницаемой в покое для ионов калия она становится избирательно проницаемой для ионов натрия. Увеличение натриевой проницаемости мембраны связано с открыванием потенциал-зависимых натриевых каналов.
Механизм, который обеспечивает открывание и закрывание ионных каналов, получил название ворот канала. Принято различать активационные (m) и инактивационные (h) ворота. Ионный канал может находиться в трех основных состояниях: закрытом (m-ворота закрыты; h-открыты), открытом (m- и h-ворота открыты) и инак-тивированном (m-ворота открыты, h- ворота закрыты) (рис 1.16).
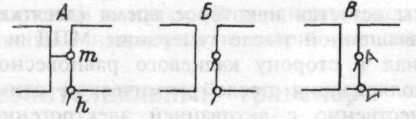
Рис. 1.16 Схема положения активационных (m) и инактивационных (h) ворот натриевых каналов, соответствующие закрытому (покой, А), открытому (активация, Б) и инактивированному (В) состояниям.
Деполяризация мембраны, вызываемая раздражающим стимулом, например, электрическим током, открывает m-ворота натриевых каналов (переход из состояния А в Б) и обеспечивает появление направленного внутрь потока положительных зарядов — ионов на-
5/
трия. Это ведет к дальнейшей деполяризации мембраны, что, в свою очередь, увеличивает число открытых натриевых каналов и, следовательно, повышает натриевую проницаемость мембраны. Возникает "регенеративная" деполяризация мембраны, в результате которой потенциал внутренней стороны мембраны стремится достичь величины натриевого равновесного потенциала.
Причиной прекращения роста МПД и реполяризации мембраны клетки является: а) увеличение деполяризации мембраны, т.е. когда Ем -» ENa, в результате чего снижается электрохимический градиент для ионов натрия, равный Ем —> ENa. Другими словами, уменьшается сила, "толкающая" натрий внутрь клетки; б) деполяризация мембраны порождает процесс инактивации натриевых каналов (закрывание h-ворот; состояние В канала), который тормозит рост натриевой проницаемости мембраны и ведет к ее снижению; в) деполяризация мембраны увеличивает ее проницаемость для ионов калия. Выходящий калиевый ток стремится сместить мембранный потенциал в сторону калиевого равновесного потенциала.
Снижение электрохимического потенциала для ионов натрия и инактивация натриевых каналов уменьшает величину входящего натриевого тока. В определенный момент времени величина входящего тока натрия сравнивается с возросшим выходящим током — рост МПД прекращается. Когда суммарный выходящий ток превышает входящий, начинается реполяризация мембраны, которая также имеет регенеративный характер. Начавшаяся реполяризация ведет к закрыванию активационных ворот (m), что уменьшает натриевую проницаемость мембраны, ускоряет реполяризацию, а последняя увеличивает число закрытых каналов и т.д.
Фаза реполяризации МПД в некоторых клетках (например, в кар-диомиоцитах и ряде гладкомышечных клеток) может замедляться, формируя плато ПД, обусловленное сложными изменениями во времени входящих и выходящих токов через мембрану. В последействии МПД может возникнуть гиперполяризация или/и деполяризация мембраны. Это так называемые следовые потенциалы. Следовая гиперполяризация имеет двоякую природу: ионную и метаболическую. Первая связана с тем, что калиевая проницаемость в нервном волокне мембраны остается некоторое время (десятки и даже сотни миллисекунд) повышенной после генерации МПД и смещает мембранный потенциал в сторону калиевого равновесного потенциала. Следовая гиперполяризация после ритмической стимуляции клеток связана преимущественно с активацией электрогенного натриевого насоса, вследствие накопления ионов натрия в клетке.
Причиной деполяризации, развивающейся после генерации МПД, является накопление ионов калия у наружной поверхности мембраны. Последнее, как это следует из уравнения Гольдмана, ведет к увеличению МПП.
С инактивацией натриевых каналов связано важное свойство нервного волокна, называемое рефрактерностью. Во время абсолютного рефрактерного периода нервное волокно полностью утрачивает способность возбуждаться при действии раздражителя любой
52
силы. Относительная рефрактерность, следующая за абсолютной, характеризуется более высоким порогом возникновения МПД.
Представление о мембранных процессах, происходящих во время возбуждения нервного волокна, служит базой для понимания и явления аккомодации. В основе аккомодации ткани при малой крутизне нарастания раздражающего тока лежит повышение порога возбуждения, опережающее медленную деполяризацию мембраны. Повышение порога возбуждения почти целиком определяется инактивацией натриевых каналов. Роль повышения калиевой проницаемости мембраны в развитии аккомодации состоит в том, что оно приводит к падению сопротивления мембраны. Вследствие снижения сопротивления скорость деполяризации мембраны становится еще медленнее. Скорость аккомодации тем выше, чем большее число натриевых каналов при потенциале покоя находится в инактивиро-ванном состоянии, чем выше скорость развития инактивации и чем выше калиевая проницаемость мембраны.
Проведение возбуждения. Проведение возбуждения по нервному волокну осуществляется за счет локальных токов между возбужденным и покоящимися участками мембраны. Последовательность событий в этом случае представляется в следующем виде.
При нанесении точечного раздражения на нервное волокно в соответствующем участке мембраны возникает потенциал действия. Внутренняя сторона мембраны в данной точке оказывается заряженной положительно по отношению к соседней, покоящейся. Между точками волокна, имеющими различный потенциал, возникает ток (локальный ток), направленный от возбужденного (знак (+) на внутренней стороне мембраны) к невозбужденному (знак (-) на внутренней стороне мембраны) к участку волокна. Этот ток оказывает деполяризующее влияние на мембрану волокна в покоящемся участке и при достижении критического уровня деполяризации мембраны в данном участке возникает МПД. Этот процесс последовательно распространяется по всем участкам нервного волокна.
В некоторых клетках (нейронах, гладких мышцах) МПД имеет не натриевую природу, а обусловлен входом ионов Ca2+ по потенциал-зависимым кальциевым каналам. В кардиомиоцитах генерация МПД связана с входящими натриевым и натрий-кальциевым токами.