

2 курс / Нормальная физиология / Физиология дыхания (2)
.pdfФИЗИОЛОГИЯ ДЫХАНИЯ.
1.Дыхание, его основные этапы. Механизмы внешнего дыхания. Биомеханика вдоха и выдоха.
Дыхание – сложный непрерывный процесс, в результате которого постоянно обновляется газовый состав крови.
В процессе дыхания различают три звена: внешнее, или легочное дыхание, транспорт газов кровью и внутреннее, или тканевое, дыхание.
Дыхание — совокупность физиологических процессов, обеспечивающих непрерывное поступление кислорода к тканям, использование его в окислительных реакциях, а также удаление из организма образующихся в процессе метаболизма углекислого газа и частично воды. К системе органов дыхания относятся носовая полость, гортань, бронхи и легкие. Дыхание состоит из следующих основных этапов:
•внешнего дыхания, обеспечивающего газообмен между легкими и внешней средой;
•газообмена между альвеолярным воздухом и притекающей к легким венозной кровью;
•транспорта газов кровью;
•газообмена между артериальной кровью и тканями;
•тканевого дыхания.
Внешнее дыхание — это газообмен между организмом и окружающим его атмосферным воздухом. Осуществляется в два этапа — обмен газов между атмосферным и альвеолярным воздухом и газообмен между кровью легочных капилляров и альвеолярным воздухом. Аппарат внешнего дыхания включает в себя дыхательные пути, легкие, плевру, скелет грудной клетки и ее мышцы, а также диафрагму. Основной функцией аппарата внешнего дыхания является обеспечение организма кислородом и освобождение его от избытка углекислого газа. О функциональном состоянии аппарата внешнего дыхания можно судить по ритму, глубине, частоте дыхания, по величине легочных объемов, по показателям поглощения кислорода и выделения углекислого газа и т. д.
Транспорт газов осуществляется кровью. Он обеспечивается разностью парциального давления (напряжения) газов по пути их следования: кислорода от легких к тканям, углекислого газа от клеток к легким.

Внутреннее или тканевое дыхание также может быть разделено на два этапа. Первый этап - обмен газов между кровью и тканями. Второй — потребление кислорода клетками и выделение ими углекислого газа (клеточное дыхание).
Вдох и выдох Вдох начинается с сокращения дыхательных (респираторных) мышц.
Мышцы, сокращение которых приводит к увеличению объема грудной полости, называются инспираторными, а мышцы, сокращение которых приводит к уменьшению объема грудной полости, называются экспираторными. Основной инспираторной мышцей является мышца диафрагмы. Сокращение мышцы диафрагмы приводит к тому, что купол ее уплощается, внутренние органы оттесняются вниз, что приводит к увеличению объема грудной полости в вертикальном направлении.
Сокращение наружных межреберных и межхрящевых мышц приводит к увеличению объема грудной полости. Легкие покрыты серозной оболочкой - плеврой, состоящей
из висцерального и париетального листков. Париетальный листок соединен с грудной клеткой, а висцеральный - с тканью легких. При увеличении объема грудной клетки, в результате сокращения инспираторных мышц, париетальный листок последует за грудной клеткой. В результате появления адгезивных сил между листками плевры, висцеральный листок последует за париетальным, а вслед за ними и легкие. Это приводит к возрастанию отрицательного давления в плевральной полости и к увеличению объема легких, что сопровождается снижением в них давления, оно становится ниже атмосферного и воздух начинает поступать в легкие - происходит вдох.
Между висцеральным и париетальным листками плевры находится щелевидное пространство, которое называется плевральной полостью. Давление в плевральной полости всегда ниже атмосферного, его называют отрицательным давлением. Величина отрицательного давления в плевральной полости равна: к концу максимального выдоха - 1-2 мм рт. ст., к концу спокойного выдоха - 2-3 мм рт. ст., к концу спокойного вдоха -5-7 мм рт. ст., к концу максимального вдоха - 15-20 мм рт. ст.

Отрицательное давление в плевральной полости обусловлено так называемой эластической тягой легких - силой, с которой легкие постоянно стремятся уменьшить свой объем. Эластическая тяга легких обусловлена двумя причинами:
•наличием в стенке альвеол большого количества эластических волокон;
•поверхностным натяжением пленки жидкости, которой покрыта внутренняя поверхность стенок альвеол.
Вещество, покрывающее внутреннюю поверхность альвеол называется сурфактантом. Сурфактант имеет низкое поверхностное натяжение и стабилизирует состояние альвеол, а именно, при вдохе он предохраняет альвеолы от перерастяжения (молекулы сурфактанта расположены далеко друг от друга, что сопровождается повышением величины поверхностного натяжения), а при выдохе - от спадения (молекулы сурфактанта расположены близко друг к другу, что сопровождается снижением величины поверхностного натяжения).
Значение отрицательного давления в плевральной полости в акте вдоха проявляется при поступлении воздуха в плевральную полость, т. е. пневмотораксе. Если в плевральную полость поступает небольшое количество воздуха, легкие частично спадаются, но вентиляция их продолжается. Такое состояние называется закрытым пневмотораксом. Через некоторое время воздух из плевральной полости всасывается и легкие расправляются.
При нарушении герметичности плевральной полости, например, при проникающих ранениях грудной клетки или при разрыве ткани легкого в результате его поражения каким-либо заболеванием, плевральная полость сообщается с атмосферой и давление в ней становится равным атмосферному, легкие спадаются полностью, их вентиляция прекращается. Такой пневмоторакс называется открытым. Открытый двусторонний пневмоторакс несовместим с жизнью.
Частичный искусственный закрытый пневмоторакс (введение в
плевральную полость с помощью иглы некоторого количества воздуха) применяется с лечебной целью, например, при туберкулезе частичное спадение пораженного легкого способствует заживлению патологических полостей (каверн).
При глубоком дыхании в акте вдоха участвуют ряд вспомогательных дыхательных мышц, к которым относятся: мышцы шеи, груди, спины.

Сокращение этих мышц вызывает перемещение ребер, что оказывает содействие инспираторным мышцам.
При спокойном дыхании вдох осуществляется активно, а выдох пассивно. Силы, обеспечивающие спокойный выдох:
•сила тяжести грудной клетки;
•эластическая тяга легких;
•давление органов брюшной полости;
•эластическая тяга перекрученных во время вдоха реберных хрящей. В активном выдохе принимают участие внутренние межреберные мышцы, задняя нижняя зубчатая мышца, мышцы живота.
2.Современные представления о структуре и локализации дыхательного центра. Автоматия дыхательного центра.
Ритмическая последовательность вдоха и выдоха, а также изменение характера дыхательных движений в зависимости от состояния организма (покой, работа различной интенсивности, эмоциональные проявления и т. д.) обусловлены наличием дыхательного центра, расположенного в продолговатом мозге Дыхательным центром называется совокупность нейронов, обеспечивающих деятельность аппарата дыхания и его приспособление к изменяющимся условиям внешней и внутренней среды.
Функциональная организация дыхательного центра.
На рисунке - нижняя часть ствола мозга (вид сзади). ПН - нейроны пневмотаксиса; ИНСП - инспираторные нейроны; ЭКСП - экспираторные нейроны.
Перерезка выше линии 1 на дыхании не отражается. Перерезка по линии 2 отделяет центр пневмотаксиса. Перерезка ниже линии 3 вызывает прекращение дыхания

Решающее значение в определении локализации дыхательного центра и его активности имели исследования отечественного физиолога Н. А. Миславского, который доказал существование дыхательного центра.
Результаты исследований Н. А. Миславского легли в основу современных представлений о локализации, строении и функции дыхательного центра. Было показано, что в дыхательном центре имеются две группы нейронов - инспираторные (вдыхательные) и экспираторные (выдыхательные). Обнаружены некоторые особенности в работе дыхательного центра. При спокойном дыхании активна только небольшая часть дыхательных нейронов и, следовательно, в дыхательном центре есть резерв нейронов, который используется при повышенной потребности организма в кислороде. Установлено, что между инспираторными и экспираторными нейронами дыхательного центра существуют функциональные взаимосвязи. Они выражаются в том, что при возбуждении инспираторных нейронов, обеспечивающих фазу вдоха, деятельность экспираторных нервных клеток заторможена и наоборот. Таким образом, одной из причин ритмичной, автоматической деятельности дыхательного центра являются взаимосвязанные функциональные отношения между вдыхательными и выдыхательными нейронами. Дыхательный центр обладает автоматией. В нем автоматически возникает ритмическое возбуждение с частотой 14 - 16 раз в минуту вследствие обменных процессов в самих нейронах, которые очень чувствительны к недостатку кислорода. Поскольку дыхательная мускулатура образована поперечно-полосатой мышечной тканью, человек способен произвольно изменять вентиляцию легких. Это возможно потому, что деятельность дыхательного центра продолговатого мозга находится под контролем коры больших полушарий.
Автоматия нейронов объясняется ритмическими колебаниями обменных процессов в них, а также воздействием на них углекислого газа. Эфферентные пути от бульбарного дыхательного центра идут к мотонейронам дыхательных межреберных и диафрагмальных мышц. Мотонейроны диафрагмальных мышц находятся в передних рогах 3-4 шейных сегментов спинного мозга, а межреберных в передних рогах грудных сегментов. Вследствие этого перерезка на уровне 1-2 шейных сегментов ведет к прекращению сокращений дыхательных

мышц. В передней части варолиева моста также имеются группы нейронов участвующих в регуляции дыхания. Кроме этих отделов ЦНС в регуляции дыхания участвуют гипоталамус, лимбическая система, кора больших полушарий. Они осуществляют более тонкую регуляцию дыхания.
3. Газообмен в легких и тканях. Основные закономерности перехода газов через мембрану. Парциальное давление и напряжение газов.
Во время вентиляции воздуха в легких происходит изменение химического состава и физических свойств поступающего в них атмосферного воздуха. В сухом воздухе при температуре 0° С и давлении 760 мм рт. ст., выдыхаемом взрослым человеком при спокойном дыхании, содержится 16,4% кислорода, 4,1% углекислого
газа и 79,5% азота. Однако при температуре 37° С альвеолярный воздух насыщен водяными п а р а м и, д а в л е н и е ко т о р ы х п р и э т о й
температуре составляет 50 мм рт. ст. Поэтому давление газов в альвеолярном воздухе равно 710 мм (760—50), содержание в нем кислорода 14—14,5%, углекислого газа 5,3—6% и азота 80—
80,5%.
Для газообмена между альвеолярным воздухом и венозной кровью, притекающей в капилляры легких, имеет значение разница в них парциальных давлений кислорода и углекислого газа. Парциальное давление кислорода, или та часть давления, которая приходится на его долю из общего давления альвеолярного воздуха, составляет 102— 110 мм рт. ст., а в венозной крови 37— 40 мм рт. ст. Вследствие этой разницы давлений в 70 мм рт. ст. кислород диффундирует из альвеолярного воздуха через стенки альвеол и капилляров в венозную кровь, превращая ее в артериальную. Парциальное давление углекислого газа в венозной крови 47 мм рт. ст., а в альвеолярном воздухе — 40 мм рт. ст. Вследствие этой разницы давления в 7 мм рт. ст. углекислый газ диффундирует из венозной крови в альвеолярный воздух и удаляется из организма при выдохе. Благодаря изменениям частоты и глубины дыхания парциальное давление углекислого газа в альвеолярном воздухе относительно постоянно, а парциальное давление кислорода в альвеолярном воздухе уменьшается пропорционально падению его парциального
давления во вдыхаемом воздухе, н а п р и м е р, п р и п о д ъ е м е н а большую высоту. Для сохранения жизни человека достаточно разности парциального давления к и с л о р о д а в а л ь в е о л я р н о м воздухе и венозной крови в несколько мм рт. ст., а углекислого газа — в 0,03 мм.
В капиллярах тканей кислород из а р т е р и а л ь н о й к р о в и диффундирует через их стенки и мембраны клеток внутрь клеток и в о в н е к л е т о ч н о е в е щ е с т в о
благодаря разности давления в 100 мм рт. ст. и больше, так как в результате обмена веществ давление кислорода в тканях доходит до нуля. А давление углекислого газа в тканях в результате обмена веществ повышается до 60—70 мм рт. ст.
Поэтому углекислый газ диффундирует через мембраны клеток и стенки капилляров в венозную кровь, где его давление равно 47 мм рт. ст.
Транспорт газов. Кислород, поглощаемый венозной кровью в капиллярах легких, соединяется с восстановленным гемоглобином и переносится артериальной кровью в ткани в виде оксигемоглобина. Все виды связей гемоглобина подробно указаны в теме «Физиология крови»
Диффузия газов через аэрогематический барьер В организме газообмен О2 и СО2 через альвеолярно-капиллярную
мембрану происходит с помощью диффузии. Диффузия О2 и СО2 через аэрогематический барьер зависит от следующих факторов: вентиляции дыхательных путей; смешивания и диффузии газов в альвеолярных протоках и альвеолах; смешивания и диффузии газов через аэрогематический барьер, мембрану эритроцитов и плазму альвеолярных капилляров; химической реакции газов с различными компонентами крови, и наконец от перфузии кровью легочных капилляров.
Диффузия газов через альвеолярно-капиллярную мембрану легких осуществляется в два этапа. На первом этапе диффузионный перенос

газов происходит по концентрационному градиенту через тонкий аэрогематический барьер, на втором — происходит связывание газов в крови легочных капилляров. После преодоления аэрогематического барьера газы диффундируют через плазму крови в эритроциты. Значительным препятствием на пути диффузии О2 является мембрана эритроцитов. Плазма крови практически не препятствует диффузии газов в отличие от альвеолярно-капиллярной мембраны и мембраны эритроцитов. Особенности диффузии газов через аэрогематический барьер количественно характеризуются через диффузионную способность легких.
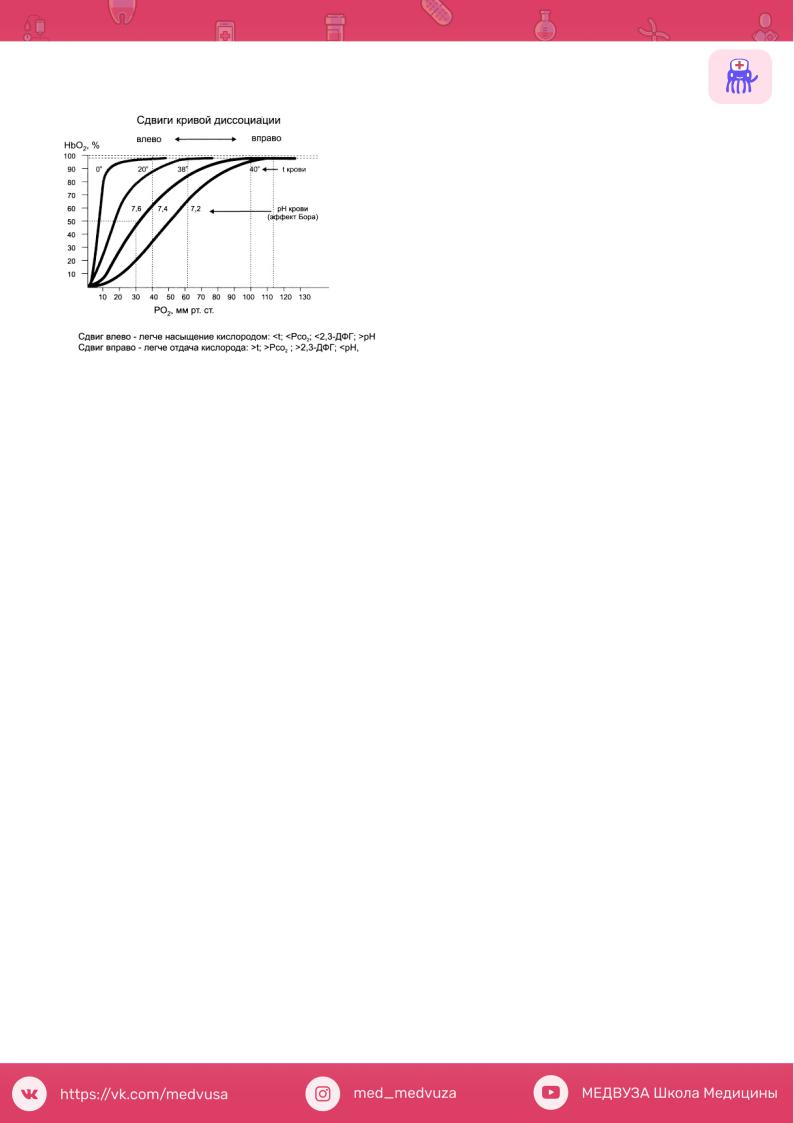
4. Кривая диссоциации оксигемоглобина, ее характеристика. Кислородная емкость крови.
Кислородная емкость крови - максимальное количество кислорода, обратимо связанное кровью; выражается в объемных процентах; зависит от концентрации в крови гемоглобина. Кислородная емкость крови человека ок. 18-20%.
Кривая диссоциации оксигемоглобина
В исходной точке гемоглобин не содержит кислорода и напряжение кислорода в крови также равняется нулю. По мере повышения напряжения гемоглобин начинает быстро насыщаться кислородом, превращаясь в оксигемоглобин:
небольшого увеличения напряжения кислорода оказывается достаточно д л я с у щ е с т в е н н о г о п р и р о с т а содержания НЬО2. При 40 мм рт. ст.
содержание НЬО2 достигает уже 75 %. Затем наклон кривой становится все более и более пологим. На этом участке кривой гемоглобин уже менее охотно присоединяет к себе кислород,
и для насыщения оставшихся 25 % НЬ требуется поднять напряжение с 40 до 150 мм рт. ст. Впрочем, в естественных условиях гемоглобин артериальной Крови никогда не насыщается кислородом полностью.
5. Рефлекторно-гуморальные механизмы регуляции дыхания. Механизм первого вдоха новорожденного.

Специфическим регулятором активности нейронов дыхательного центра является углекислый газ, который действует на дыхательные нейроны непосредственно и опосредованно. В ретикулярной формации продолговатого мозга, вблизи дыхательного центра, а
также в области сонных синусов и дуги аорты обнаружены хеморецепторы, чувствительные к углекислому газу. При увеличении напряжения углекислого газа в крови хеморецепторы возбуждаются, и нервные импульсы поступают к инспираторным нейронам, что приводит к повышению их активности.
Углекислый газ повышает возбудимость нейронов коры головного мозга. В свою очередь клетки КГМ стимулируют активность нейронов дыхательного центра.
При оптимальном содержании в крови углекислого газа и кислорода наблюдаются дыхательные движения, отражающие умеренную степень возбуждения нейронов дыхательного центра. Эти дыхательные движения грудной клетки получили название эйпноэ. Избыточное содержание углекислого газа и недостаток кислорода в крови усиливают активность дыхательного центра, что обусловливает возникновение частых и глубоких дыхательных движений –гиперпноэ. Еще большее нарастание количества углекислого газа в крови приводит к нарушению ритма дыхания и появлению одышки – диспноэ.
Механизм первого вдоха новорожденного.
В организме матери газообмен плода происходит через пупочные сосуды. После рождения ребенка и отделения плаценты указанная связь нарушается. Метаболические процессы в организме новорожденного приводят к образованию и накоплению углекислого газа, который, так же как и недостаток кислорода, гуморально возбуждает дыхательный центр. Кроме того, изменение условий существования ребенка приводит к возбуждению экстеро- и проприорецепторов, что также является одним из механизмов, принимающих участие в осуществлении первого вдоха новорожденного.
Рефлекторные механизмы.
Различают постоянные и непостоянные (эпизодические) рефлекторные влияния на функциональное состояние дыхательного центра.

Постоянные рефлекторные влияния возникают в результате раздражения рецепторов альвеол (рефлекс Геринга — Брейера), корня легкого и плевры (пульмоторакальный рефлекс), хеморецепторов дуги
аорты и сонных синусов (рефлекс Гейманса), проприорецепторов дыхательных мышц.
Рефлекс Геринга — Брейера является одним из механизмов саморегуляции дыхательного процесса, обеспечивая смену актов вдоха и выдоха. При растяжении альвеол во время вдоха нервные импульсы от рецепторов растяжения по блуждающему нерву идут к экспираторным нейронам, которые, возбуждаясь, тормозят активность инспираторных нейронов, что приводит к пассивному выдоху. Легочные альвеолы спадаются, и нервные импульсы от рецепторов растяжения уже не поступают к экспираторным нейронам. Активность их падает, что создает условия для повышения возбудимости инспираторной части дыхательного центра и осуществлению активного вдоха.
Кроме того, активность инспираторных нейронов усиливается при нарастании концентрации углекислого газа в крови, что также способствует проявлению вдоха.
Пульмоторакальный рефлекс возникает при возбуждении рецепторов, заложенных в легочной ткани и плевре. Проявляется этот рефлекс при растяжении легких и плевры. Рефлекторная дуга замыкается на уровне шейных и грудных сегментов спинного мозга. К дыхательному центру постоянно поступают нервные импульсы от проприорецепторов дыхательных мышц. Во время вдоха происходит возбуждение проприорецепторов дыхательных мышц и нервные импульсы от них поступают в инспираторную часть дыхательного центра. Под влиянием нервных импульсов тормозится активность вдыхательных нейронов, что способствует наступлению выдоха. Непостоянные рефлекторные влияния на активность дыхательных нейронов связаны с возбуждением разнообразных экстеро- и интерорецепторов
6. Дыхание в условиях пониженного и повышенного барометрического давления.
Снижение парциального давления кислорода в окружающем воздухе, что в естественных условиях имеет место при восхождении человека