

2 курс / Нормальная физиология / ФЗЛ МЫШ И НЕРВОВ
.pdfФИЗИОЛОГИЯ МЫШЦ И НЕРВОВ
Структура миофибрилл и ее изменения при сокращении. Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнах миофибриллы разделены на правильно чередующиеся участки (диски), обладающие разными оптическими свойствами. Одни участки анизотропны, т. е. обладают двойным лучепреломлением. В обыкновенном свете они выглядят темными, а. в поляризованном— прозрачными в продольном направлении й непрозрачными в поперечном. Другие участки в Обыкновенной свете выглядят светлыми — они изотропны, т. е. не обладают двойным лучепреломлением. Анизотропные участки обозначают буквой А, изотропные — буквой I. В середине диска А различается светлая полоска Н, посередине диска / — темная полоска -представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы. Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрилл внутри одного волокна во время сокращения не смещаются по отношению друг к другу.
Молекула актина Это глобулярный белок, состоящий из одного полипептида, который
полимеризуется с другими молекулами актина и образует две цепи, обвивающие друг друга. Такая двойная спираль представляет собой остов тонкого филамента. На каждой молекуле актина есть участок связывания миозина. В покоящемся
мышечном волокне взаимодействие между актином и миозином предотвращают два белка - тропонин и тропомиозин.
Тропомиозин - стержневидная молекула из двух полипептидов, обвивающихся друг около друга; молекула соответствует в длину примерно семи мономерам актина. Цепи из молекул тропомиозина, уложенные конец в конец, располагаются вдоль всего тонкого филамента. Молекулы тропомиозина частично прикрывают участки связывания каждой молекулы актина, мешая контакту миозина с актином. В таком блокирующем положении молекула тропомиозина удерживается тропонином.
Тропонин - гетеротримерный белок. Он состоит из тропонина Т (отвечает з а с в я з ы в а н и е с о д и н о ч н о й
молекулой тропомиозина), тропонина С (связывает ион Са 2+) и тропонина I (связывает актин и ингибирует сокращение). Каждая молекула тропомиозина связана с одной гетеротримерной молекулой тропонина, которая регулирует доступ к участкам связывания миозина на семи мономерах актина, прилегающих к молекуле тропомиозина.
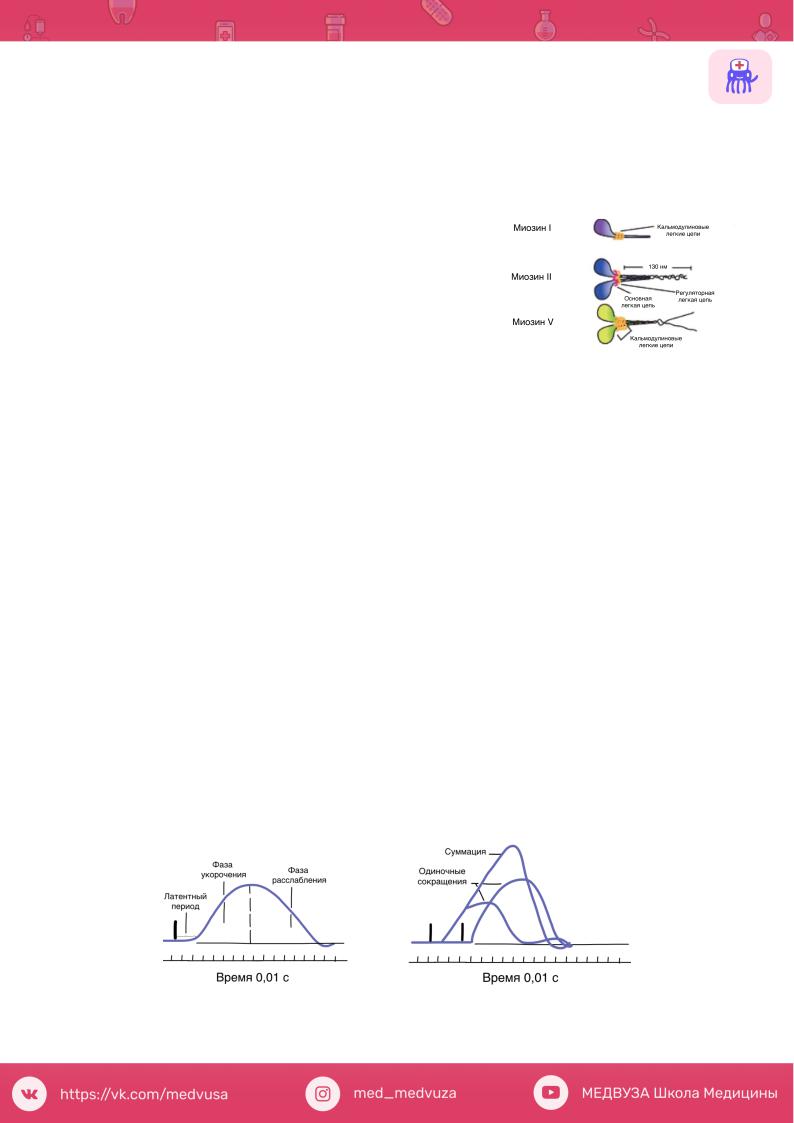
Миозин Это единое название большой семьи протеинов, имеющих определенные
отличия в клетках разных тканей. Миозин присутствует у всех эукариотов.
В последнее время показано, что миозин |
II участвует в мышечном |
|
сокращении, тогда как миозин |
I и миозин |
V вовлечены в работу |
подмембранного (кортикального) цитоскелета. В настоящее время идентифицировано более 10 классов миозина.
На рисунке показаны схемы строения миозина, который состоит из головки, шейки и хвоста. Молекула миозина состоит из двух больших полипептидов (тяжелых цепей) и четырех меньших (легких цепей). Эти полипептиды составляют молекулу с двумя глобулярными «головками», которые содержат оба вида цепей, и длинным стержнем («хвостом») из двух
переплетенных тяжелых цепей. Хвост каждой молекулы миозина располагается вдоль оси толстого филамента, а две глобулярные головки выступают по бокам, их иначе называют поперечными мостиками. На каждой глобулярной головке находятся по два участка связывания: для актина и для АТФ. Участки связывания АТФ обладают также свойствами фермента АТФазы, гидролизующей связанную молекулу АТФ.
Суммация сокращений и ее виды. Тетанус и его виды. Оптимум и пессимум.
Мышечное сокращение является жизненно важной функцией организма, связанной с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Все виды произвольных движений осуществляются за счет скелетных мышц.
Непроизвольные движения (кроме сокращения сердца) обусловлены сокращением гладких мышц. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Одиночное мышечное сокращение. При раздражении мышцы одиночным импульсом тока возникает одиночное мышечное сокращение.
Амплитуда
Амплитуда одиночного сокращения мышцы зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных групп волокон различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна. При увеличении силы раздражающего тока в процесс возбуждения вовлекаются и менее возбудимые группы мышечных волокон; амплитуда сокращений суммируется и растет до тех пор, пока в мышце не останется волокон, не охваченных процессом возбуждения. В этом случае регистрируется максимальная амплитуда сокращения, которая не увеличивается, несмотря на дальнейшее нарастание силы раздражающего тока.
Фазы и режимы сокращения скелетной мышцы
Фазы мышечного сокращения При раздражении скелетной мышцы одиночным импульсом электрического тока
сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы:
•латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения;
•возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
•фаза укорочения (около 50 мс);
•фаза расслабления (около 50 мс).
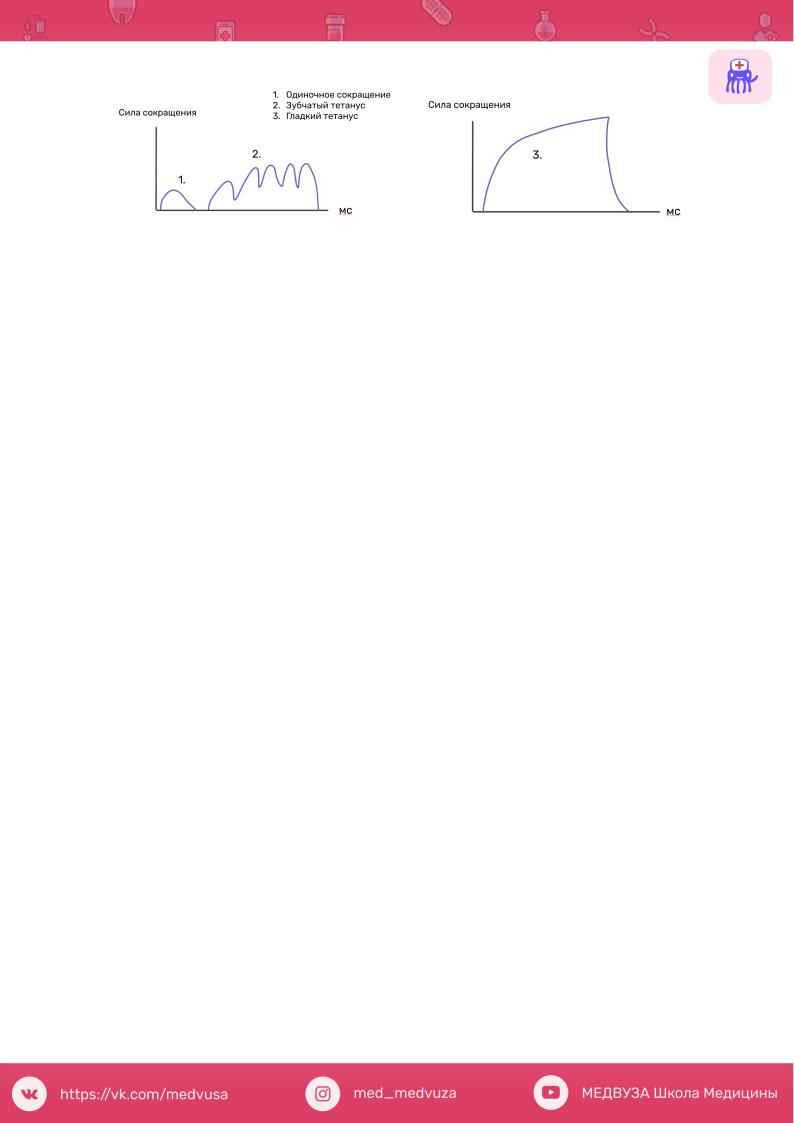
Тетанус, тетаническое мышечное сокращение — состояние длительного сокращения, непрерывного напряжения мышцы, возникающее при поступлении к ней через мотонейрон нервных импульсов с высокой частотой. При этом расслабления между последовательными одиночными сокращениями не происходит и возникает их суммация, приводящая к стойкому максимальному сокращению мышцы.
Виды тетануса, механизм возникновения различных видов тетануса:
Если последующее раздражение наносится, когда мышца уже начала расслабляться после предыдущего укорочения (то есть приходится на фазу
расслабления предыдущего цикла) -наблюдается зубчатый тетанус (неполный).

На миографической кривой вершина второго сокращения будет отделена от вершины первого небольшим западением кривой (рис. ниже).
Если последующее раздражение приходится на фазу укорочения предыдущего цикла, наблюдается гладкий тетанус (полный). На миографической кривой второе сокращение полностью сливается с первым, образуя единую вершину.
В порядке возрастания амплитуды сокращения можно расставить режимы сокращения так: одиночное мышечное сокращение, зубчатый тетанус, гладкий тетанус, Причем амплитуда зубчатого и гладкого тетануса зависит от частоты раздражения. Во время одиночного сокращения в поперечных мостиках между нитями актина и миозина возникает упругое напряжение, однако одиночного стимула недостаточно для прикрепления всех мостиков. Высокий уровень Са2+обеспечивает образование большего количества поперечных мостиков, что в свою очередь повышает амплитуду укорочения мышцы. При этом чем больше частота стимуляции, тем выше амплитуда укорочения, но до определенного предела.
В латентный период мышечного сокращения происходят последовательно следующие процессы:
•распространение потенциала действия по сарколемме и системе поперечных трубочек вглубь мышечного волокна;
•возбуждение мембран цистерн саркоплазматического ретикулума и открытие кальциевых каналов;
•выход из концевых цистерн ионов кальция;
•диффузия ионов кальция в межфибриллярное пространство;
•освобождение активных участков актиновых нитей для связывания с головкой миозина.
Наименьшая частота стимуляции мышцы, при которой амплитуда гладкого тетануса максимальная, называется оптимум частоты раздражения. При дальнейшем повышении частоты стимуляции происходит расслабление мышцы, такая частота получила название пессимальной. Уменьшение частоты стимуляции тотчас же приводит к восстановлению исходного высокого уровня тетанического сокращения
Оптимум силы |
- сила раздражителя, при действии которого возбуждением |
охвачены все мышечные волокна и амплитуда тетануса максимальна. |
|
Пессимум силы |
- сила раздражителя, превышающая оптимальную, которая |
вызывает уменьшение амплитуды тетануса вплоть до полного расслабления мышцы.
В целостном организме мышцы сокращаются в режимах:
-одиночного сокращения и зубчатого тетануса, характерных для медленных ДЕ;
-“ложного” тетануса -то есть в виде ряда последовательных одиночных
сокращений, свойственных быстрым ДЕ.
Однако форма сокращения целостной мышцы напоминает гладкий тетанус.
Причина этого -асинхронность разрядов мотонейронов и сократительной
реакции отдельных мышечных волокон (мышечные волокна, относящиеся к одной двигательной единице, сокращаются синхронно). Благодаря этому мышца плавно сокращается и плавно расслабляется, а также может длительно находиться в

сокращенном состоянии за счет чередования сокращений множества мышечных волокон.
Классификация синапсов. Строение синапса.Механизм передачи возбуждения в синапсах
1. По местоположению и принадлежности соответствующим структурам:
Øпериферические (нервно-мышечные, нейросекреторные, рецепторнонейрональные);
Øцентральные (аксо-соматические, аксо-дендритные, аксоаксональные, сомато-дендритные. сомато-соматические);
2. По эффекту действия:
Øвозбуждающие
Øтормозные
3. По способу передачи сигналов:
Øэлектрические. Передают возбуждение без участия медиатора с большой скоростью и обладают двухсторонним проведением возбуждения. Структурной основой электрического синапса является нексус. Встречаются эти синапсы в железах внутренней секреции, эпителиальной ткани, ЦНС, сердце.
Øхимические,
Øсмешанные.
Внекоторых органах возбуждение может передаваться и через химические и через электрические синапсы.
4) По медиатору, с помощью которого осуществляется передача сигнала химические синапсы классифицируют:
Øхолинергические,
Øадренергические,
Øсеротонинергические,
Øглицинергически. и т.д.
Синапсы с химической передачей возбуждения обладают рядом общих свойств:
Ø Возбуждение через синапсы проводится только в одном направлении (односторонне). Это обусловлено строением синапса: медиатор выделяется только из пресинаптического утолщения и взаимодействует с рецепторами постсинаптической мембраны;

Øпередача возбуждения через синапсы осуществляется медленнее, чем по нервному волокну — синаптическая задержка;
Øпередача возбуждения осуществляется с помощью специальных химических посредников — медиаторов;
Øв синапсах происходит трансформация ритма возбуждения;
Øсинапсы обладают низкой лабильностью;
Øсинапсы обладают высокой утомляемостью;
Øсинапсы обладают высокой чувствительностью к химическим (в том числе и к фармакологическим: блокаторам, психомиметикам).
ПД достигая нервного окончания (пресинаптической мембраны) вызывает его деполяризацию. При деполяризации пресинаптической мембраны кальций входит в пресинаптическую терминаль через специфические потенциалозависимые кальциевые каналы в этой мембране. Увеличение концентрации кальция в нервном окончании способствует освобождению ацетилхолина, который выходит в синаптическую щель. Медиатор достигает постсинаптической мембраны и связывается там с рецепторами. В результате внутрь постсинаптической мембраны поступают ионы натрия и эта мембрана частично деполяризуется, т.е. возбуждение пока еще не распространяется дальше, а находится в синапсе. Частичная деполяризация постсинаптической мембраны называется возбуждающим постсинаптическим потенциалом (ВПСП).
Врезультате этих механизмов развивается синаптическая задержка, которая составляет от 0,2 до 1 мВ.
Под влиянием ВПСП в соседнем чувствительном участке мембраны мышечного волокна возникает распространяющийся ПД, который и вызывает сокращение мышцы.
Для восстановления возбудимости постсинаптической мембраны после очередного импульса необходима инактивация медиатора (инактивационная система). В противном случае, при длительном действии медиатора происходит снижение чувствительности рецепторов к этому медиатору.
Для заблокирования передачи возбуждения через синапс применяют яд кураре, который связывается с рецепторами постсинаптической мембраны и препятствует их взаимодействию с ацетилхолином.
Заблокировать проведение возбуждения через синапс может яд бутулин и другие вещества.
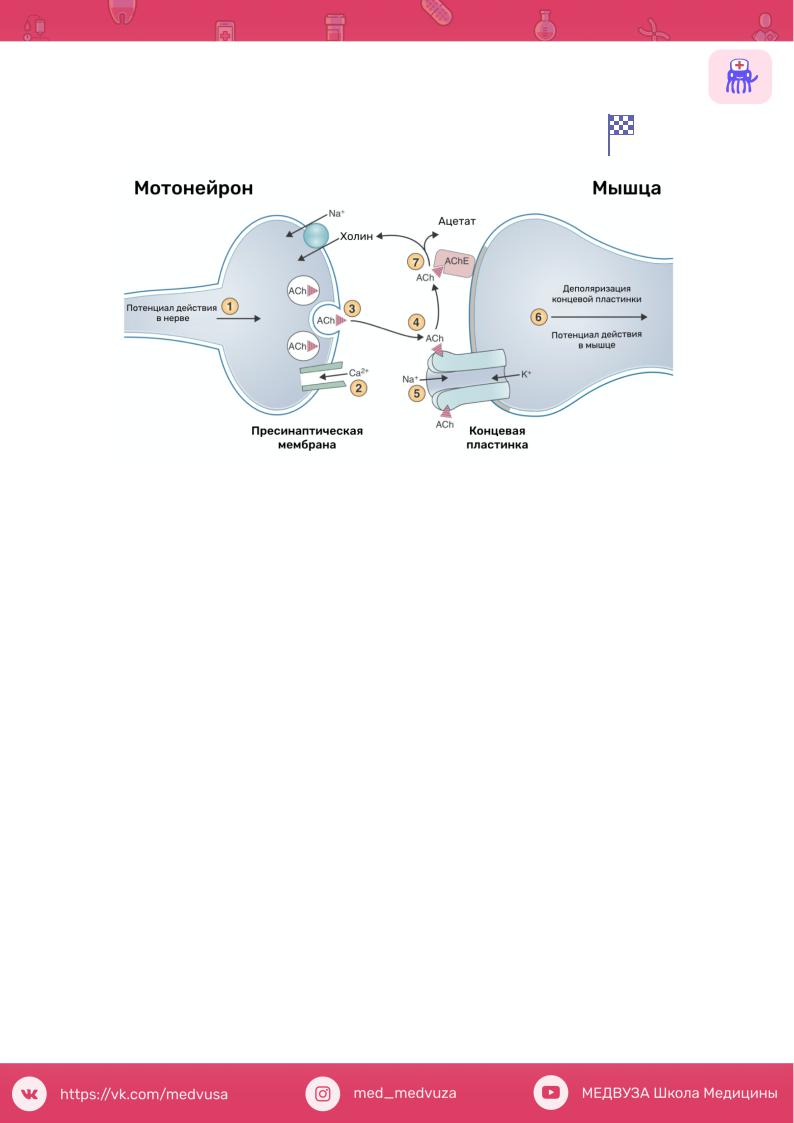
Механизм синаптической передачи возбуждения.
Советую тут интенсивно работать со скриптом, ниже предложен дополнительный вариант механизма передачи
1.Деполяризация (возбуждение) пресинаптической мембраны.
2.Изменение проницаемости для ионов кальция.
3.Ионы кальция или его ионизированные комплексы по концентрационному градиенту поступают в нервное окончание (антагонистами кальция являются ионы магния и токсины ботулинуса).
4.Уменьшение электростатических влияний (одноименных зарядов) между пресинаптической мембраной и везикулами.
5.Приближение и слияние везикул с пресинаптической мембраной.
6.Изменение поверхностного натяжения везикул.
7.Разрыв везикул.
8.Выход медиатора в синоптическую щель.
9.Медиатор (возбуждающий в нервно-мышечном синапсе: ацетилхолин) диффундирует через синоптическую щель к рецепторам постсинаптической мембраны.
10.Ацетилхолин вступает во взаимодействие с холинорецепторами (обладают избирательной чувствительностью к ацетилхолину).
11.При одновременном участии ионов кальция и макроэргического фосфата происходят конформационные изменения белковых молекул рецептора.
12.Открываются каналы постсинаптической мембраны для Na или Са.
13.Ионы Na по концентрационному градиенту поступают внутрь воспринимающей возбуждение клетки.
14.Развивается деполяризация – возбуждающий постсинаптический потенциал, который носит местный характер, по форме и свойствам напоминает локальный ответ (не подчиняется закону «все или ничего» и способен суммироваться).
15.Суммация возбуждающих постсинаптических потенциалов
16.Потенциал концевой пластинки.
17.Когда он достигает определенной (критической величины) возникают местные токи между возбужденными участками постсинаптической мембраны и невозбужденными участками прилегающей к ней обычной (электровозбудимой) мембраной.

18. На прилегающем участке электровозбудимой мембраны возникает потенциал действия.
Классификация нервных волокон. Распространения возбуждения по безмиелиновым и миелиновым нервным волокнам. Характеристика их возбудимости и лабильности. Законы проведения возбуждения по нерву.
Наиболее распространена классификация по Дж.Эрлангеру и Х.Гассеру (1937), в которой волокна разделяют на три типа: А, В и С. Волокна типа А и В являются миелиновыми, типа С — безмиелиновыми. Волокна А делят на 4 подгруппы: α, β, γ, δ. В периферической нервной системе к
волокнам А α относятся афферентные волокна от механорецепторов кожи, мышечных и сухожильных рецепторов, а также эфферентные волокна к скелетным мышцам. К А β принадлежат афферентные волокна от кожных рецепторов прикосновения и давления, от части мышечных и
висцеральных рецепторов. А γ представляют собой эфферентные волокна, через которые регулируется активность мышечных рецепторов. К А δ относят афферентные волокна от части тактильных, температурных и болевых, а также суставных рецепторов. К волокнам типа В принадлежат преганглионарные волокна вегетативной нервной системы. К волокнам типа С относят постганглионарные волокна вегетативной нервной системы, афферентные волокна от некоторых б о л е в ы х (вторичная боль), т е п л о в ы х и в и с ц е р а л ь н ы х рецепторов.
И з д а н н ы х, представленных в таблице, видно, ч т о с р е д н и й диаметр каждого т и п а в о л о к н а снижается от типа
А до С (каждый примерно в 2 раза по отношению к предыдущему). Соответственно этому снижается и скорость проведения возбуждения. Низкая скорость проведения нервного импульса в волокнах типа С связана с особенностями проведения возбуждения в безмиелиновых
волокнах. Лабильность также уменьшается от волокон А α до С и находится в обратной зависимости от продолжительности фазы абсолютной рефрактерности. Возбудимость тоже уменьшается от волокон А α (наибольшая возбудимость) к волокнам С (наименьшая

возбудимость). Например, пороговая сила электрического тока у волокон С в 30—50 раз больше, чем у волокон А α. Исследование факторов, блокирующих нервную проводимость, показало, что к давлению наиболее чувствительны волокна А, к кислородному голоданию (гипоксии) — волокна В, к местным анестетикам — волокна С.
Немиелинизированные волокна.
Проведение ПД по ННВ происходит путем активации потенциалзависимых натриевых ионных каналов участка мембраны волокна, прилегающего к тому месту, где возник ПД. При этом между возбужденным и невозбужденным участками мембраны нервного волокна возникают локальные электрические токи, в ы з ы в а ю щ и е д е п о л я р и з а ц и ю м е м б р а н ы невозбужденного участка до критического уровня. После этого в мембране НВ мгновенно открываются потенциалзависимые натриевые каналы. ПД, т.о., генерируется в последующем участке НВ.
Миелинизированные волокна.
В МНВ ПД генерируются только в области перехвата Ранвье, т.е. той части мембраны, которая не покрыта шванновскими клетками. С одной стороны, это обусловлено тем, что цитоплазма шванновской клетки содержит липид –
сфингомиелин, который уменьшает поток ионов через мембрану НВ примерно в 5000 раз и снижает
ее емкость в 50 раз. С другой стороны, в области ПР в мембране НВ имеется наибольшее число потенциалзависимых натриевых ионных каналов, а сама мембрана обладает нормальным уровнем возбудимости. При нанесении точечного удара на МНВ ПД генерируется в зоне ПР, и возникают электрические токи, которые текут вдоль силовых линий от плюса к минусу потенциала на мембране. Одновременно в аксоплазме НВ возникают продольные токи, которые направлены от места
генерации ПД в обе стороны. Продольные токи вызывают открывание ПЗНИК, а следовательно, движение ионов Na+ через мембрану и генерацию ПД в соседних ПР, минуя часть НВ, покрытого шванновскими клетками. Поскольку ПД передаются (перепрыгивают) от одного ПР к другому, то механизм проведения ПД в МНВ получил название сальтаторного. В МНВ реполяризация мембраны после ПД происходит с очень высокой скоростью. В результате НВ имеют высокую функциональную лабильность и способны проводить значительное число ПД в единицу времени. Скорость распространения ПД в зависимости от диаметра и типа МНВ чрезвычайно высока. В толстых МНВ скорость проведения возбуждения пропорциональна диаметру волокна, а проводимость НВ имеет обратную зависимость от его диаметра.

Законы проведения возбуждения по нервному волокну:
1. Закон физиологической непрерывности – проведение возбуждения по НВ
возможно при условии его структурной целостности и физиологической непрерывности. Физиологическая непрерывность нерва м.б. нарушена, например, при сдавлении нерва без его структурного повреждения, что препятствует проведению ПД.
2. Закон изолированного проведения – при проведении возбуждения по НВ ПД
не распространяется с одного волокна на другое, например, рядом расположенное.
3. Закон двухстороннего проведения – отдельно нервное волокно обладает
двухсторонней проводимостью. Так, при искусственном электрическом раздражении в любой точке по ходу НВ может возникать ПД и распространяться как центростремительно, так и центробежно.
Лабильность. Парабиоз и его фазы (Н.Е.Введенский).
Лабильность — функциональная подвижность, скорость протекания элементарных циклов возбуждения в нервной и мышечной тканях. Понятие "Л." введено русским физиологом Н. Е. Введенским (1886), который считал мерой Л. наибольшую частоту раздражения ткани, воспроизводимую ею без преобразования ритма. Лабильность отражает время, в течение которого ткань восстанавливает работоспособность после очередного цикла возбуждения. Наибольшей лабильностью отличаются отростки нервных клеток — аксоны, способные воспроизводить до 500—1000 импульсов в 1 сек; менее лабильны центральные и периферические места контакта — синапсы (например, двигательное нервное окончание может передать на скелетную мышцу не более 100—150 возбуждений в 1 сек). Угнетение жизнедеятельности тканей и клеток (например, холодом, наркотиками) уменьшает лабильность, т. к. при этом замедляются процессы восстановления и удлиняется рефрактерный период. Парабиоз — состояние, пограничное между жизнью и смертью клетки.
Причины парабиоза – самые разные повреждающие воздействия на возбудимую ткань или клетку, не приводящие к грубым структурным изменениям, но в той или иной мере нарушающее ее функциональное состояние. Такими причинами могут быть механические, термические, химические и другие раздражители.
Сущность парабиоза. Как считал сам Введенский, в основе парабиоза лежит снижение возбудимости и проводимости, связанное с натриевой инактивацией. Советский цитофизиолог Н.А. Петрошин полагал, что в основе парабиоза лежат обратимые изменения белков протоплазмы. Под действием повреждающего агента клетка (ткань), не теряя структурной целостности, полностью прекращает функционировать. Это состояние развивается фазно, по мере действия повреждающего фактора (то есть зависит от продолжительности и силы действующего раздражителя). Если повреждающий агент вовремя не убрать, то наступает биологическая смерть клетки (ткани). Если же этот агент убрать вовремя, то ткань так же фазно возвращается в нормальное состояние.
Эксперименты Н.Е. Введенского.