

6 курс / Гастроэнтерология / Клинические_перспективы_гастроэнтерологии,_гепатологии_2004_№05
.pdfНаучно0
практический журнал для клиницистов
№ 5, 2004
:
%.'. ( #
:
).). *
:
+.,. -* ( ), 8.(. -* , +.%. 9 , '.8. 8 , +.;. 8 , <.%. < , +.%. , , +.=. ' * , +.+. & *
:
> ,
,,, «( «<-% »
:
,,, «( «<-% »
: 10 000 .
:
1 2
:
B * * «-2004», I:
41727 – * ;
41728 –
82127 – * «) . E* »«> » 2- * 2004 .
E* < >; , ! * 30.06.2000 .
( ( № 77-3872)
:
125284, < , / 74 E-mail: rm-vesti@mtu-net.ru
*( - http://www.m-vesti.ru
#
, * *
© «9 , », 2004
Российская гастроэнтерологическая ассоциация Российское общество по изучению печени
Содержание
Маевская М.В.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
Ешану В.С.
. . . . . . . . . . . . . . . . . . . 11
Буеверов А.О., Маевская М.В.
o. . . . . . . . . . . . . . . . . . . . . . . . . . 17
Козлова Н.М., Тюрюмин Я.Л., Кулинский В.И., Леонова З.А., Якобсон Ю.А.
! . . . . . . . . . . . 23
Пасечников В.Д., Минушкин О.Н., Алексеенко С.А., Котелевец С.М., Мостовов А.Н., Чуков С.З., Масловский Л.В., Зверьков И.В., Володин Д.В.
" Helicobacter pylori# ?
( ) . . . . . . . . . 27
Калинин А.В., Логинов А.Ф., Дзюба К.В., Белоусов Е.Л. $ « »
: . . . . . . . . . . 31
% . . . . . . . . . . . . . . . . 35
& . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36

Гастроэнтерология, базирующаяся на доказательствах
Клинические перспективы гастроэнтерологии, гепатологии ■ 5, 2004
УДК 616.36 004.4 056.7
Патогенез алкогольной болезни печени и роль генетической предрасположенности в ее развитии
<.%. <
( . . . )
Системное окисление этанола происходит в печени по трем метаболическим путям: параллельно с использованием алкогольдегидрогеназы (АДГ), микросомальной этаноло0 окислительной системы и каталаз пироксисом. Систематическое воздействие алкоголя на0 рушает множество разнообразных метаболических путей. Однако, что же обусловливает реализацию алкогольной болезни печени (АБП), еще не ясно. Активно изучается роль гене0 тической предрасположенности в развитии АБП, в частности полиморфизм генов АДГ, аль0 дегиддегидрогеназы, цитохрома Р450 2Е1. Результаты исследований противоречивы, а са0 ма проблема требует дальнейшего изучения.
Ключевые слова: алкогольная болезнь печени, генетическая предрасположенность.
П |
атогенез |
алкогольной |
тия фиброза. Активно изучается |
хождения этанола в желудке. При |
|||
болезни |
печени |
(АБП) |
роль |
апоптоза в |
алкогольном |
быстром сокращении желудка, на0 |
|
изучается |
четыре |
деся0 |
повреждении печени. Благодаря |
пример, натощак, первый этап ме0 |
|||
тилетия. Известно, что |
появлению новых идей создаются |
таболизма этанола уменьшается. |
|||||
действие |
алкоголя нарушает мно0 |
новые направления научных иссле0 |
При медленном опорожнении же0 |
||||
жество разнообразных метаболи0 |
дований и лечения АБП. |
лудка (после еды) контакт этанола |
|||||
ческих путей в печени. Однако не0 |
В слизистой оболочке желудка |
с его слизистой оболочкой удлиня0 |
|||||
понятно, что же наиболее важно в |
осуществляется первый этап мета0 |
ется, что приводит к метаболизму |
|||||
реализации АБП как таковой. По0 |
болизма алкоголя |
при участии |
большей части алкоголя. |
||||
этому предсказать индивидуальный |
желудочной фракции алкогольде0 |
В то же время вопрос о желуд0 |
|||||
риск развития АБП пока невоз0 |
гидрогеназы (АДГ): γ0АДГ, χ0АДГ, |
ке как основном органе, где осу0 |
|||||
можно. |
|
|
|
δ0АДГ (рис. 1). Подробная характе0 |
ществляется первая фаза метабо0 |
||
В последние 5 лет появились но0 |
ристика АДГ приводится ниже. |
лизма этанола, дискутируется. Вы0 |
|||||
вые концепции алкогольного пора0 |
Активность желудочной АДГ зави0 |
сказывается предположение, что |
|||||
жения печени. Жировая печень |
сит от пола и возраста пациента, |
первая фаза метаболизма этано0 |
|||||
рассматривается как предраспо0 |
приема им лекарств, а также, воз0 |
ла происходит в печени [8], одна0 |
|||||
лагающий фактор развития более |
можно, от колонизации желудка |
ко на нее влияет скорость абсорб0 |
|||||
тяжелого повреждения; звездчатым |
Heliсobacter pylori [8, 14]. |
ции этанола из желудка и тонкой |
|||||
ретикулоэндотелиоцитам (клеткам |
Первый этап метаболизма ал0 |
кишки. |
|||||
Купфера) отводится главная роль в |
коголя |
имеет большое значение, |
Системное окисление этанола |
||||
координации воспалительного от0 |
так как от этого зависит его коли0 |
происходит в печени параллельно |
|||||
вета на воздействие алкоголя, что |
чество, достигающее органов0ми0 |
по трем метаболическим путям: |
|||||
является |
основой развития |
алко0 |
шеней. Из самого желудка этанол |
с использованием АДГ, микросо0 |
|||
гольного |
гепатита. Стали |
более |
всасывается медленно, но при по0 |
мальной этанолоокислительной си0 |
|||
понятными факторы контроля акти0 |
ступлении в верхние отделы тонкой |
стемы (МЭОС) и каталаз пирокси0 |
|||||
вации reticuloendotheliocytus stella0 |
кишки его всасывание ускоряется. |
сом (рис. 2). |
|||||
tus как центрального звена разви0 |
Большое значение имеет время на0 |
Первые два пути (с использова0 |
|||||
2

Клинические перспективы гастроэнтерологии, гепатологии ■ 5, 2004
нием АДГ и МЭОС) изучены хоро0 шо. Они приводят к окислению эта0 нола соответственно в ацетальдегид в цитозоле и эндоплазматическом ретикулуме. Каталазы в основном содержатся в пироксисомах и могут участвовать в окислении этанола, что, однако, ограничивается доступ0 ностью необходимого им субстрата
– перекиси водорода (Н2О2). Дальнейший метаболизм ацет0
альдегида протекает с использова0 нием цитозольной (АЛДГ1) и (или) митохондриальной альдегиддегид0 рогеназы (АЛДГ2). В результате об0 разуется ацетат, большая часть ко0 торого далее окисляется вне пече0 ни. Ферменты, изоферменты и их гены, участвующие в метаболизме этанола, отличаются полиморфиз0 мом. Именно благодаря этому факту отдельные лица могут легче переносить этанол [8].
АДГ ответственны за метабо0 лизм основного количества приня0 того внутрь алкоголя. Они пред0 ставляют собой наиболее древнее семейство ферментов, имеющееся у всех позвоночных, насекомых, многих водорослей и микробов. Изоферменты АДГ обозначаются греческими буквами α, β, γ, π, χ, δ, μ. Все они представляют собой ди0 меры с субъединицами молекуляр0 ной массой 40 кДa.
Изоферменты АДГ сгруппиро0 ваны в классы на основании их ферментативных характеристик и нуклеотидных последовательнос0 тей (табл. 1).
Печеночная АДГ – цитоплазма0 тический фермент, расположенный преимущественно в центре пече0 ночной дольки. АДГ – цинкзависи0 мый фермент. Поэтому при дефи0 ците в организме цинка активность АДГ снижается с последующим за0 медлением метаболизма алкоголя. У человека существуют три основ0 ных гена, кодирующих АДГ: АДГ1, АДГ2 и АДГ3.
Локусы АДГ2 и АДГ3 полиморф0 ны. В то же время на АДГ1 локусе идентифицирован только один ал0 лель. Ферменты, закодированные на АДГ1 локусе и на полиморфном локусе АДГ3, имеют некоторое
Класс I АДГ
Класс II АДГ CYP2E1
Каталазы
Класс I АДГ
Класс IV АДГ
Рис. 1. Метаболизм этанола в желудке и печени [8, p. 97]
Таблица 1. Свойства АДГ человека [8, p. 101, с изменениями]
Локус (расположение аллельных |
Изоферменты |
Распределение |
|
генов в хромосоме) |
в тканях |
||
|
|||
|
|
|
|
Kласс I: |
α |
|
|
АДГ1 |
Печень |
||
АДГ2 |
β |
Печень, легкие |
|
АДГ3 |
γ |
Печень, желудок |
|
Kласс II: |
π |
Печень, роговица |
|
АДГ4 |
|||
Kласс III: |
χ |
|
|
АДГ5 |
Большинство тканей |
||
Kласс IV: |
δ, μ |
|
|
АДГ7 |
Cлизистые oболочки |
||
|
|
желудка, пищевода и др. |
|
Kласс V: |
|
|
|
АДГ6 |
– |
Печень, желудок |
|
Kласс VI: |
|
|
|
АДГ8 |
– |
Не выявлен у человека |
сходство по их кинетическим кон0 стантам. Для АДГ2 обнаружено 3 аллеля, для АДГ3 – 2 (табл. 2).
Полиморфизм на локусе АДГ2, вероятнее всего, приводит к значи0 тельным различиям в метаболизме этанола. Изофермент АДГβ1 (ал0 лель АДГ2*1) чаще встречается у европейцев, АДГβ2 (аллель АДГ2*2) – у лиц азиатского проис0 хождения; у европейцев частота составляет 5–20%.
У аллеля АДГ2*2 – так называе0 мого «атипичного аллеля» – актив0 ность в 3–6 раз выше, чем у других аллелей. Плохая переносимость алкоголя и такие симптомы, как по0 краснение лица, сердцебиение и т. д. (flush0синдром), у лиц азиатско0 го происхождения связывают с вы0 сокой частотой этого аллеля и ге0
нетической редукцией активности изоэнзима АЛДГ1.
АДГ0β3 (аллель АДГ2*3) в срав0 нении со всеми другими изоформа0 ми АДГ класса 1 обладает значи0 тельно большей способностью и максимальной скоростью окисле0 ния этанола. Аллель АДГ2*3 обна0 руживается у афро0американцев с частотой 15–20%.
D.G. McCaver и соавт. [17] предполагают, что этот аллель об0 ладает защитными свойствами от токсического действия алкоголя. Они изучали полиморфизм гена АДГ2 и частоту врожденных дефек0 тов у афро0американских ново0 рожденных, матери которых употребляли алкоголь до и во вре0 мя беременности. Полиморфизм АДГ2 также изучали и у матерей.
Гастроэнтерология, базирующаяся на доказательствах
3

Гастроэнтерология, базирующаяся на доказательствах
Клинические перспективы гастроэнтерологии, гепатологии ■ 5, 2004
|
|
|
|
|
|
|
|
|
|
АЛДГ2 |
||
А |
|
|
|
АДГ |
|
|
|
|
АЛДГ1 |
|||
|
Этанол |
|
|
|
|
|
Ацетальдегид |
|
|
|
Ацетат |
|
|
|
|
NAD+ |
|
|
|
|
|
|
NAD+ |
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
NADH |
|
|
NADH |
||||
|
|
|
|
|
|
|||||||
|
|
|
CYP2E1 |
|
|
|
|
|||||
Б |
|
Этанол |
|
|
|
|
|
Ацетальдегид |
|
|
|
|
|
|
|
NAD+ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
NADH |
|
|
|
|
|||
|
|
|
|
|
|
|
|
|||||
|
|
|
Каталазы |
|
|
|
|
|||||
В |
|
|
|
|
|
|
|
|
|
|
|
|
|
Этанол |
|
|
|
|
|
Ацетальдегид |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Н2O2 |
|
|
O2 |
|
|
|
|
||
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
Рис. 2. Метаболистические пути окисления этанола в печени: А – посредством АДГ, Б – посредством МЭОС (CYP2E1), В – посредством каталаз пироксисом [8, p. 10]
Таблица 2. Полиморфизм АДГ человека [8, p. 109]
Локус (расположение аллельных |
Тип |
*K (этанол) |
**V |
Распределение в популяциях |
|
генов в хромосоме) |
субъединиц |
||||
m |
max |
|
|||
|
|
|
|
|
|
АДГ2*1 |
β1 |
0,05 |
9 |
Европейцы, aфро0американцы |
|
АДГ2*2 |
β2 |
0,9 |
400 |
Азиаты |
|
АДГ2*3 |
β3 |
34 |
300 |
Афро0американцы |
|
АДГ3*1 |
γ1 |
1,0 |
87 |
Все популяции |
|
АДГ3*2 |
γ2 |
0,63 |
35 |
Европейцы |
* Km – константа Михаэлиса , единица измерения – мМ.
** Vmax – максимальная скорость ферментативной химической реакции, достигаемая постепенным увеличением концентрации субстрата (единица измерения – мин–1).
По результатам проведенного |
болизма этанола. Например, ал0 |
ной специфичности к алифатиче0 |
|||
исследования ментальный статус |
лель АДГ2*3 увеличивает скорость |
ским и ароматическим альдегидам. |
|||
новорожденных не |
отличался от |
окисления этанола на 10% [29]. |
Классификация АЛДГ основана на |
||
такового в группе контроля (непью0 |
Однако это не совсем просто, |
их электрофоретической подвиж0 |
|||
щие женщины) при наличии аллеля |
так как фермент АДГ метаболизи0 |
ности и не вполне стандартизова0 |
|||
АДГ2*3 как у пьющей матери, так и |
рует не весь алкоголь, а только |
на (табл. 3). |
|
||
у ребенка. Отсутствие этого алле0 |
часть его, поступившего в орга0 |
Основное участие в метабо0 |
|||
ля у пьющих матерей и их новорож0 |
низм. Более того, скорость его |
лизме этанола |
принимают цито0 |
||
денных сочеталось со сниженным |
окисления варьирует у лиц с одина0 |
зольная АЛДГ1 и митохондриаль0 |
|||
индексом умственного развития у |
ковым генотипом АДГ и даже у |
ная АЛДГ2. Оба фермента имеют |
|||
детей. Это обстоятельство дало ос0 |
близнецов [8]. |
тетрамерное строение и состоят |
|||
нование предположить, что аллель |
Меньшая вариабельность ме0 |
из субъединиц молекулярной мас0 |
|||
АДГ2*3 обладает защитными свой0 |
таболизма этанола связана с локу0 |
сой 54 кДa. Экспрессия АЛДГ1 и |
|||
ствами от токсического |
действия |
сом АДГ3. Относительно полимор0 |
АЛДГ2 наблюдается не только в |
||
алкоголя. |
|
|
физма АДГ3 существует мнение, |
печени, но и в других органах (поч0 |
|
Предполагается, что у различ0 |
что он не влияет на скорость эли0 |
ки, мышцы, сердце, поджелудочная |
|||
ных индивидуумов комбинации изо0 |
минации алкоголя [5]. |
железа). АЛДГ5 имеет нуклеотид0 |
|||
энзимов АДГ2 и АДГ3 с вариабель0 |
Окисление этанола АДГ приво0 |
ную последовательность, близкую |
|||
ностью Vmax и Km для этанола обус0 |
дит к продукции ацетальдегида – |
АЛДГ2. Какую роль в метаболизме |
|||
ловливают различную |
скорость |
высокотоксичного соединения, ко0 |
ацетальдегида |
играют АЛДГ6–9, |
|
его окисления. |
|
|
торое метаболизируется преиму0 |
неизвестно [8]. |
|
Таким образом, |
по |
присутст0 |
щественно никотинамидаденинди0 |
Окисление |
ацетальдегида по0 |
вию определенного |
аллеля АДГ |
нуклеотидзависимыми АЛДГ. Эти |
средством АЛДГ требует участия |
||
можно предсказать скорость мета0 |
ферменты имеют спектр субстрат0 |
никотинамидадениндинуклеотида |
|||
4
|
|
|
|
|
|
|
|
Клинические перспективы гастроэнтерологии, гепатологии |
■ 5, 2004 |
|
|||||
|
|
NADPH + H+ + O |
2 |
|
NADP+ + 2H O |
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
2 |
|
|
|
|
|
доказательствах |
||
Алкоголь |
|
|
|
М Э О С |
Ацетальдегид |
|
Ацетат |
|
|
||||||
|
|
|
Гладкий эндоплазматический |
|
|
|
|
|
|||||||
|
|
|
|
|
ретикулум |
|
|
|
|
|
|
||||
|
|
Алкогольдегидрогеназа |
Альдегиддегидрогеназа |
Ацетил4КоА |
|||||||||||
|
Цитозоль |
NAD+ |
|
NADH + H+ |
NAD+ |
NADH + H+ |
Цикл Кребса |
||||||||
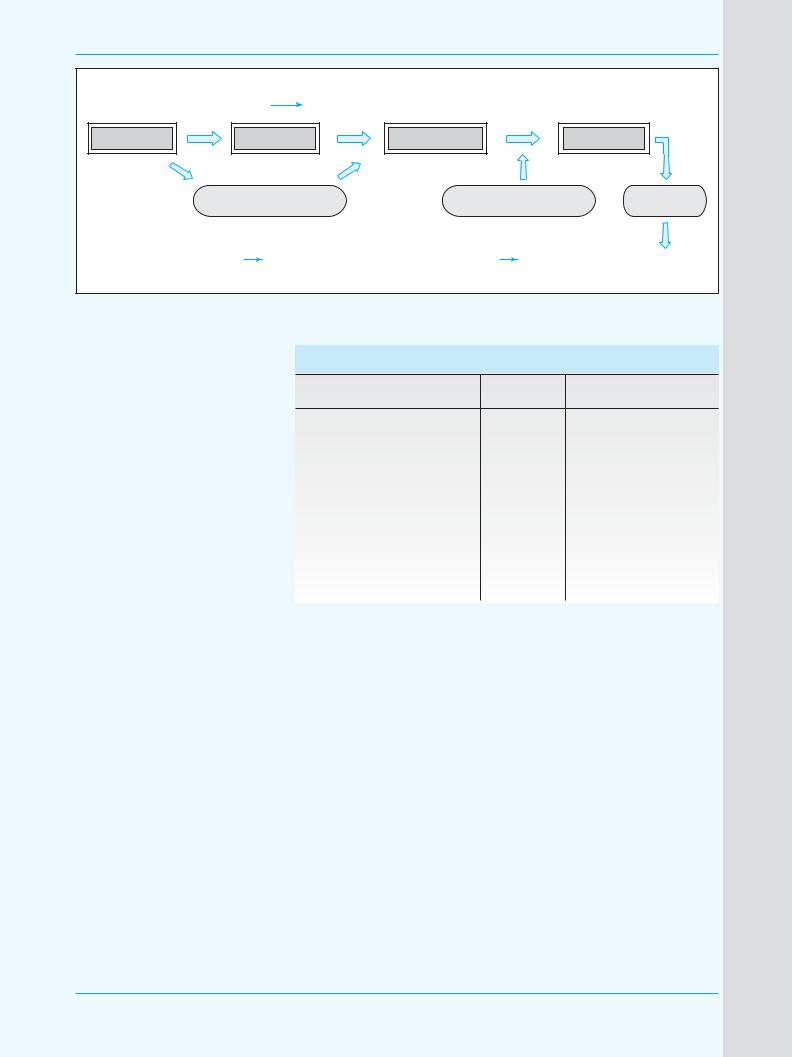
Рис. 3. Метаболические пути окисления этанола (по E. Kuntz, H.4D. Kuntz, 2002 [14] |
|
|
|
||||||||||||
(NAD), что сопровождается пере0 |
Таблица 3. Свойства АЛДГ человека [8, p. 106, с изменениями] |
||||||||||||||
носом атома водорода с молекулы |
|
||||||||||||||
|
|
|
|
|
|
на |
|||||||||
этанола |
на |
молекулу |
NAD |
с |
Локус (расположение аллельных |
Структура |
Распределение |
||||||||
его восстановлением |
до |
NADH |
|||||||||||||
генов в хромосоме) |
в тканях |
|
|||||||||||||
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
(рис. 3). Далее высокотоксичный |
Kласс I: |
|
α4 |
|
|
|
базирующаяся |
||||||||
ацетальдегид также в NAD0зависи0 |
АЛДГ1 |
|
Различные ткани |
|
|||||||||||
мой реакции превращается в мито0 |
|
|
|
Печень > почки |
|
||||||||||
Kласс 2: |
|
|
|
|
|
||||||||||
хондриях с помощью АЛДГ в уксус0 |
|
α4 |
Печень > почки > мышцы |
||||||||||||
АЛДГ2 |
|
||||||||||||||
ную кислоту. Образующийся при |
|
||||||||||||||
|
|
|
> сердце; в большинстве |
||||||||||||
этом NADH повторно окисляется. |
|
АЛДГ5 |
|
? |
тканей уровень низкий |
||||||||||
Скорость деградации алкоголя |
|
Печень > почки > мышцы |
|||||||||||||
|
|
|
> сердце; в большинстве |
||||||||||||
с помощью АДГ зависит от интен0 |
|
|
|
||||||||||||
|
|
|
тканей уровень низкий |
||||||||||||
сивности |
повторного |
окисления |
Kласс 3: |
|
α2 |
|
|
|
|||||||
NADH в NAD. Метаболизм одной |
АЛДГ3 |
|
Желудок, печень, роговица |
||||||||||||
молекулы алкоголя требует две мо0 |
Другие ферменты: |
|
α4 |
Печень |
|
|
|||||||||
лекулы NAD + H+. Это значительно |
АЛДГ9 |
|
|
|
|||||||||||
АЛДГ6–8 |
|
? |
? |
|
|
||||||||||
изменяет |
окислительно0восстано0 |
|
|
|
|||||||||||
|
|
|
|
|
|
||||||||||
вительный |
потенциал |
печеночных |
|
|
|
|
|
|
|||||||
клеток в сторону восстановитель0 |
остановить процесс |
деградации |
ется АДЛГ2*1, а мутантный – |
||||||||||||
|
|||||||||||||||
ных реакций. Деградация алкоголя |
алкоголя. |
|
АЛДГ2*2. Дефицит АЛДГ2 переда0 |
Гастроэнтерология, |
|||||||||||
может прекратиться, если NADH |
Хорошо известно, что употреб0 |
ется как доминантный признак во |
|||||||||||||
не будет постоянно реокисляться. |
|
ление алкоголя у определенных лиц |
всех изучаемых популяциях. Среди |
||||||||||||
Повторное |
окисление |
NADH |
приводит к развитию flush0синдро0 |
печеночной АЛДГ 40% общей ак0 |
|||||||||||
происходит в |
митохондриальной |
ма. Наиболее часто это встречает0 |
тивности фермента приходится на |
||||||||||||
дыхательной цепи и заключается в |
ся у японцев, китайцев и корейцев |
АЛДГ2, а остальные 60% – на дру0 |
|||||||||||||
удалении полученного от этанола |
даже после приема небольшого ко0 |
гие формы (АЛДГ1, АЛДГ3 и, воз0 |
|||||||||||||
атома водорода с образованием |
личества алкоголя, в то время как у |
можно, АЛДГ5) [8]. |
|
|
|||||||||||
воды и выделением энергии. Об0 |
европейцев этот феномен наблю0 |
Исследования в |
этой |
области |
|||||||||||
разующаяся |
уксусная |
кислота |
дается редко. Эта реакция основа0 |
ограничены побочными эффектами |
|||||||||||
поступает в кровь и далее во вне0 |
на на аккумуляции ацетальдегида, |
алкоголя, а их результаты противо0 |
|||||||||||||
печеночных тканях при |
участии |
концентрация которого чаще всего |
речивы. |
|
|
|
|||||||||
ацетил0коэнзим А синтетазы пре0 |
превышает 100 мкM. |
|
При |
хроническом употребле0 |
|||||||||||
вращается в ацетил0КоА, который |
По данным японских авторов, у |
нии алкоголь метаболизируется |
|||||||||||||
поступает в цикл Кребса, или цикл |
лиц с flush0синдромом имеется де0 |
преимущественно |
ферментатив0 |
||||||||||||
жирных кислот, используется в син0 |
фицит АЛДГ2. Это связано с мута0 |
ной системой цитохрома Р450 IIE1 |
|||||||||||||
тезе холестерола либо окисляется |
цией гена и заменой гуанина на |
(CYP2E1), который впервые описал |
|||||||||||||
до углекислого газа (рис. 3). |
|
|
|
аденин, что приводит к замещению |
Ч. Либер в 1994 г. как МЭОС. |
||||||||||
Угнетение |
митохондриальной |
глутамата лизином в позиции 487 |
МЭОС состоит из CYP2E1 ре0 |
||||||||||||
дыхательной цепи специфическими |
полипептида АЛДГ2. |
|
дуктаз, расположенных в мембра0 |
|
|||||||||||
ингибиторами |
может |
полностью |
Нормальный аллель обознача0 |
не эндоплазматического ретикулу0 |
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
5 |
|
|

Гастроэнтерология, базирующаяся на доказательствах
Клинические перспективы гастроэнтерологии, гепатологии ■ 5, 2004
Закрашенные участки – экзоны, незакрашенные – интроны
Рис. 4. Структура гена эукариот
ма. Для метаболизма этанола эта ферментативная система исполь0 зует NADPH и О2 в качестве кофакторов и индуцируется хрони0 ческим введением алкоголя, что может увеличивать массу фермен0 та до 10 раз. Этанол играет роль донора для восстановления О2 до воды, а сам превращается в ацет0 альдегид (рис. 3).
Ген CYP2E1 имеет 11 kb в длину и разделен на 9 экзонов (см. ниже), его промотерная активность обна0 ружена в 5’0концевом регионе. Экспрессия гена обладает ткане0 вой специфичностью с наиболь0 шим его содержанием в печени преимущественно в 30й зоне аци0 нуса. В меньших количествах ген CYP2E1 экспрессируется в легких, пищеводе и кишечнике [8].
Представляется, что большие концентрации этанола повышают количество мРНК CYP2E1 за счет увеличения транскрипции гена CYP2E1. Более того, уровень мРНК CYP2E1 в мононуклеарных клетках периферической крови предлага0 ется в качестве чувствительного маркера употребления алкоголя, поскольку нормализация показате0 ля наблюдается в течение 3–4 дней на фоне абстиненции [24, 33].
CYP2E1 можно разделить на три типа: А – гомозиготы по с1 гену, В – гетерозиготы по с1 и с2 генам и С – гомозиготы по с2 гену на осно0 вании различных продуктов, кото0 рые получаются при воздействии эндонуклеаз (фосфодиэстераз).
Эндонуклеазы – ферменты, расщепляющие фосфодиэфирные связи нуклеиновых кислот с обра0 зованием поли0 и олигонуклеоти0 дов различной длины.
Рестриктирующие эндонуклеа0 зы – ферменты, расщепляющие
специфические последовательнос0 ти двухцепочечной ДНК (использу0 ются при проведении генетическо0 го анализа).
У здоровых людей преоблада0 ют генотипы А и В [3, 34]. Поли0 морфизм обнаружен в 5’0регионе гена Р450 2Е1, что может нару0 шать его транскрипционную актив0 ность. Генетические исследования количества и характера мутаций проводились по длине фрагментов ДНК – restriction length fragment polymorphism (RELF), что означает существование аллельных форм, обнаруживаемых по длине фраг0 ментов после того, как нуклеотид0 ную цепь обработали специфиче0 скими ферментами рестрикции (рестриктирующими эндонуклеаза0 ми), расщепившими ее.
В биоптатах тканей лиц, недав0 но употреблявших алкоголь, выяв0 лена четырехкратная индукция CYP2E1, что влечет за собой тяже0 лые метаболические дефекты. CYP2E1 – высококонсервативный фермент, обладающий уникальной способностью метаболизировать ксенобиотики. Одни из них при этом детоксицируются, в то время как другие превращаются в высо0 котоксичные метаболиты.
К ксенобиотикам относятся индустриальные сольвенты, анес0 тетики (например, галотан), часто используемые медикаменты (изо0 ниазид, фенилбутазон, парацета0 мол). Именно с индукцией CYP2E1 связана повышенная чувствитель0 ность злоупотребляющих алкого0 лем лиц к этим агентам.
Следует отметить, что алкого0 лики особенно подвержены токси0 ческому действию парацетамола в момент прекращения приема ал0 коголя, то есть в период абстинен0
ции, когда они испытывают разно0 образные неприятные ощущения (головную боль, диспепсию и т. д.) и особенно нуждаются в аналгези0 рующих препаратах. В этот период алкоголь, парацетамол и голода0 ние действуют как синергисты, уве0 личивая токсичность каждого ве0 щества в отдельности. Все они уменьшают запасы глутатиона, обеспечивающего одну из фунда0 ментальных клеточных функций – удаление свободных радикалов.
Индукция CYP2E1 ассоциирует0 ся с пролиферацией эндоплазма0 тического ретикулума, что сопро0 вождается повышенным окислени0 ем NADPH, продукцией свободных радикалов (Н2О2, супероксида и т. д.), что приводит к перекисному окислению липидов и, как следст0 вие, к повреждению печени.
Пероксидация липидов корре0 лирует с количеством CYP2E1 в препаратах микросом печени и по0 давляется антителами к CYP2E1 у кормленных этанолом крыс и у жи0 вотных группы контроля [15].
CYP2E1 также был обнаружен в звездчатых ретикулоэндотелиоци0 тах. При кормлении этанолом крыс в этих клетках в 7 раз повысилось содержание CYP2E1, что дополни0 тельно к эндотоксинам стимулиро0 вало их к высвобождению провос0 палительных и фиброгенных цито0 кинов [12].
При нормальных обстоятель0 ствах механизмы клеточной за0 щиты от активных радикалов пре0 дотвращают их повреждающее действие. Однако при болезнях печени, дефиците питания или максимальной индукции CYP2E1 вредные эффекты могут превали0 ровать, тогда баланс адаптивной системы нарушается.
Оба ферментативных пути – АДГ и МЭОС – генерируют токси0 ческий метаболит этанола – аце0 тальдегид, обладающий способно0 стью образования белковых ком0 плексов. Это приводит к активации ферментов, уменьшению репара0 ции ДНК, продукции антител, исто0 щению глутатиона, митохондри0 альной токсичности, нарушению
6
Клинические перспективы гастроэнтерологии, гепатологии ■ 5, 2004
|
|
|
АДГ |
|
|
|
АЛДГ2 |
|
|
|
|
с низкой активностью |
|
|
с |
высокой активностью |
|
|
|
|
|
|
|
|
|
|
|
|
Этанол |
|
|
Ацетальдегид |
|
|
Ацетат |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
АДГ, CYP2E1 |
|
|
|
АЛДГ2 |
|
|
|
|
с высокой активностью |
|
|
с высокой активностью |
||
|
|
|
|
|
|
|
|
|
|
|
Этанол |
|
|
Ацетальдегид |
|
|
Ацетат |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
АДГ |
|
|
|
АЛДГ2 |
|
|
|
|
с низкой активностью |
|
|
с низкой активностью |
||
|
|
Этанол |
|
|
Ацетальдегид |
|
|
Ацетат |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
АДГ |
|
|
|
АЛДГ2 |
|
|
|
|
с высокой активностью |
|
|
|
с низкой активностью |
|
|
|
Этанол |
|
|
Ацетальдегид |
|
|
Ацетат |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
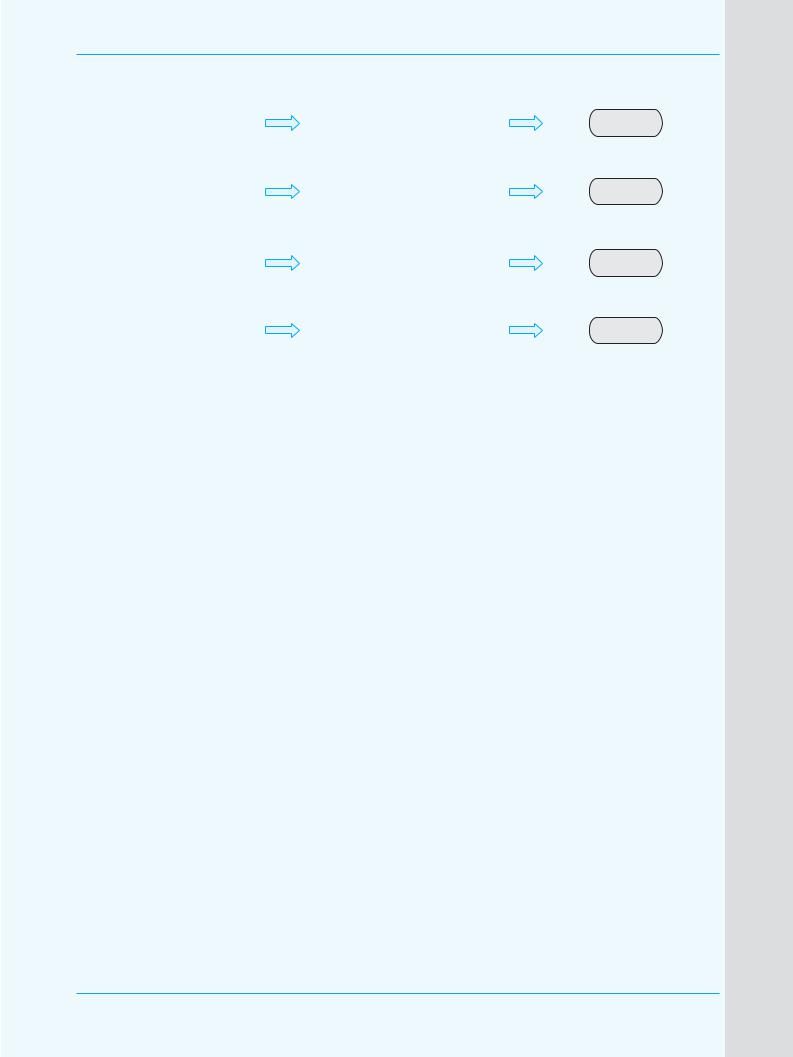
Рис. 5. Потенциальные механизмы развития АБП, основанные на генетически обусловленной разной скорости элиминации алкоголя [8, р. 112, с изменениями]
утилизации кислорода и повышен0 |
чин, 30 г/сут и выше для женщин) |
последовательностями (интронами). |
|||||||||||
ному синтезу коллагена. |
вызывают повреждение |
печени. |
Следовательно, гены эукариот |
||||||||||
Другие цитохромы Р450 (1А2, |
Тем не менее тяжелые формы АБП |
представляют собой мозаику из |
|||||||||||
3А4) также вовлечены в метабо0 |
(алкогольный гепатит и цирроз пе0 |
чередующихся в определенном по0 |
|||||||||||
лизм этанола. Их индукция вносит |
чени) |
|
развиваются |
только у |
рядке экзонов и интронов (рис. 4). |
||||||||
свой «вклад» в переносимость |
15–20% алкоголиков. В связи с |
Принципиально важное свойст0 |
|||||||||||
отдельными |
индивидуумами алко0 |
этим логично предположить о гене0 |
во генетической информации – ее |
||||||||||
голя и лекарственных препаратов, |
тических |
факторах |
ее |
развития, |
способность к передаче в преде0 |
||||||||
представляющих собой микросо0 |
определяющих различия |
в |
актив0 |
лах одной клетки, от родительской |
|||||||||
мальные |
субстраты. |
Переноси0 |
ности |
|
метаболизирующих |
алко0 |
клетки к дочерним, между клетками |
||||||
мость алкоголиками психоактивных |
голь |
|
ферментативных |
систем. |
различных индивидуумов в процес0 |
||||||||
препаратов обычно связывается с |
Результаты исследований |
в этой |
се клеточного деления и размно0 |
||||||||||
адаптивными возможностями цент0 |
области противоречивы. |
|
|
|
жения. Для ДНК0содержащих орга0 |
||||||||
ральной нервной системы (ЦНС). |
|
|
|
|
|
|
|
|
|
низмов перенос генетической ин0 |
|||
Однако в дополнение к этому не0 |
|
|
Основные этапы |
|
|
|
формации связан с копированием |
||||||
обходимо рассматривать возмож0 |
|
|
|
|
|
(репликацией) ДНК или с синтезом |
|||||||
|
|
передачи генетической |
|||||||||||
ность метаболической адаптации |
|
|
молекул РНК (транскрипцией) и по0 |
||||||||||
|
|
информации |
|
|
|
|
|||||||
ЦНС, что связано со скоростью |
|
|
|
|
|
|
следующим образованием поли0 |
||||||
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|||||
клиренса |
различных |
препаратов |
|
|
Для более полного понима0 |
пептидов (трансляцией). Каждый |
|||||||
из крови. |
|
|
|
ния сути исследований необходи0 |
процесс осуществляется на основе |
||||||||
Результаты контролируемых ис0 |
мо напомнить структуру |
генов |
принципов матричности и компле0 |
||||||||||
следований показали, что хрони0 |
эукариот и основные этапы пере0 |
ментарности. |
|||||||||||
ческое введение чистого этанола в |
дачи генетической информации. |
Транскрипция (первый этап об0 |
|||||||||||
условиях |
полноценного питания |
Гены |
эукариот имеют |
мозаич0 |
щего переноса генетической ин0 |
||||||||
как на животной модели (крысы), |
ную структуру – прерывистый ха0 |
формации) заключается в биосин0 |
|||||||||||
так и у людей приводит к порази0 |
рактер |
структурно0функциональ0 |
тезе молекул РНК по программе |
||||||||||
тельно высокому клиренсу из кро0 |
ной |
организации. |
Информация |
ДНК. Суть этого процесса состоит |
|||||||||
ви этанола, |
мепробамата, фено0 |
такого гена о структуре гипотетиче0 |
в том, что информация структурно0 |
||||||||||
барбитала, |
пропранолола. По0 |
ского синтезируемого полипептида |
го гена (структурный ген кодирует |
||||||||||
добно этому выявляется усиление |
существует не в виде непрерывной |
структуру полипептидов и молекул |
|||||||||||
метаболизма антипирина, толбу0 |
нуклеотидной последовательности |
РНК) в виде нуклеотидной последо0 |
|||||||||||
тамида, варфарина, диазепама и |
определенного участка |
молекулы |
вательности участка нити ДНК пе0 |
||||||||||
рифамицина [15]. |
|
ДНК, а в форме кодирующих фраг0 |
реписывается (транскрибируется) в |
||||||||||
Доказано, что опасные дозы |
ментов (экзонов), которые прерыва0 |
нуклеотидную последовательность |
|||||||||||
алкоголя (40 г/сут и более для муж0 |
ются |
«инертными» |
нуклеотидными |
молекулы РНК. |
|||||||||
Гастроэнтерология, базирующаяся на доказательствах
7

Гастроэнтерология, базирующаяся на доказательствах
Клинические перспективы гастроэнтерологии, гепатологии ■ 5, 2004
В качестве продуктов транс0 крипции можно рассматривать все типы молекул РНК, участвующие в биосинтезе белков в клетке. К ним относятся матричные (информаци0 онные) РНК (мРНК, или иРНК),
рибосомные РНК (рРНК), транс0 портные РНК (тРНК), малые ядер0 ные РНК (мяРНК). Процесс транс0 крипции обеспечивается комплекс0 ным действием ряда ферментов и подразделяется на три основные стадии:
1)инициация – начало синтеза
РНК;
2)элонгация – удлинение поли0 нуклеотидной цепочки;
3)терминация – окончание процесса.
Инициация транскрипции зави0 сит от предварительного связыва0 ния РНК0полимеразы с узнаваемой ею короткой нуклеотидной после0 довательностью в участке молеку0 лы ДНК (промотере), расположен0 ном перед стартовой точкой струк0 турного гена, с которой начинает0 ся синтез РНК. Специфическое прочное связывание РНК0полиме0 разы с промотером позволяет на0 чать процесс расплетания молеку0 лы ДНК до стартовой точки, с кото0 рой начинается полимеризация рибонуклеотидов с использовани0 ем в качестве матрицы одноните0 вого фрагмента ДНК.
Дальнейшее расплетание ДНК структурного гена сопровождается удлинением синтезируемого поли0 рибонуклеотида – элонгацией нити РНК. Этот процесс продолжается до достижения РНК0полимеразой области терминатора, который представляет собой нуклеотидную последовательность ДНК, узнавае0 мую РНК0полимеразой при учас0 тии других белковых факторов тер0 минации. Это приводит к оконча0 нию синтеза транскрипта и отсое0 динению его от матрицы.
В случае структурных генов эу0 кариот транскрибируется вся нук0 леотидная последовательность фрагмента ДНК, содержащая эк0 зоны и интроны. При этом образу0 ется молекула мРНК, отражающая структуру всего мозаичного гена,
которую называют гетерогенной ядерной РНК, или проматричной РНК. Далее эта молекула претер0 певает процессинг, суть которого состоит в ферментативном разре0 зании первичного транскрипта (проматричной РНК) и удалением его интронных участков с воссое0 динением (сплайсингом) экзонных участков. В результате формирует0 ся зрелая мРНК, которая в даль0 нейшем транспортируется из карио0 плазмы в цитоплазму с участием специфических транспортных бел0 ков и участвует в трансляции гене0 тической информации.
Трансляция заключается в син0 тезе полипептида на рибосоме с использованием в качестве матри0 цы мРНК и также подразделяется на три основные стадии: инициа0 цию, элонгацию и терминацию. В процессе трансляции участвуют молекулы тРНК, функции которых состоят в транспортировке амино0 кислот из цитозоля к рибосомам.
Завершающий этап реализа0 ции генетической информации –
посттрансляционная модификация полипептида, которая превращает его в функционально активную мо0 лекулу [2].
Генетическая предрас положенность к алкого лизму и АБП
Проведенные к 900м годам прошлого века исследования на близнецах показали, что существу0 ет генетический компонент, обес0 печивающий как скорость элими0 нации алкоголя у разных лиц, так и предрасположенность к АБП. Вы0 сокая конкордантность (сходство анализируемых признаков у одно0 го человека или пары) частоты цир0 роза печени для монозиготных близнецов в сравнении с таковой у дизиготных предопределяло оче0 видность генетического компонен0 та среди факторов риска развития АБП. Однако большинство цирро0 зов печени встречалась у тех лю0 дей, кто был подвержен риску раз0 вития алкоголизма [10, 13].
При более позднем анализе
этих же результатов несколько из0 менились сделанные выводы в пользу того, что риск развития ал0 когольного цирроза связан с гене0 тической предрасположенностью именно к алкоголизму, а не к по0 вреждению печени [25]. Поскольку метаболизм этанола имеет ключе0 вое значение в патогенезе АБП, логично изучать полиморфизм от0 ветственных за него генов, обеспе0 чивающих разную скорость окис0 ления алкоголя у разных лиц.
Основанная идея генетических исследований заключается в том, что присутствие высокоактивной АДГ и (или) CYP2E1 и малоактивной АЛДГ повышает скорость продук0 ции и (или) снижает скорость дегра0 дации ацетальдегида, что, в свою очередь, увеличивает риск разви0 тия АБП (рис. 5).
Доказано, что аллель АДГ2*2 чаще встречается у лиц, которые не злоупотребляют алкоголем в сравнении с алкоголиками. По данным F. Tanaka и соавт. [28], ал0 лель АДГ2*1, напротив, чаще встречается у злоупотребляющих алкоголем лиц, чем у тех, кто при0 нимает его в средних и малых до0 зах. Аллель АДГ3*1 реже встреча0 ется у алкоголиков монголоидной расы, чем у непьющих, однако у ев0 ропейцев подобной зависимости не выявлено [9].
Оценить связь между риском развития АБП и различными алле0 лями АДГ сложно, а количество ис0 следований в этой области ограни0 ченно.
Японские авторы [32] указыва0 ют на повышенный риск развития АБП у гетеро0 и гомозигот АДГ2*2 при условии продолжающегося употреблении алкоголя. Этот факт подтверждается результатами двух других исследований (4, 28), пока0 завших значительно более высо0 кий риск развития цирроза печени у лиц с аллелем АДГ2*2.
На основании исследований в Великобритании [6] и Франции [7] сделано заключение о более высо0 кой частоте АДГ3 у европейцев с АБП. При объединении результа0 тов этих двух исследований показа0
8

Клинические перспективы гастроэнтерологии, гепатологии ■ 5, 2004
но, что АДГ3*1 достоверно чаще встречается у алкоголиков с цир0 розом печени, чем у таковых без цирроза [23]. В то же время этот результат опровергнут данными другого исследования [26]. А при обобщении всех трех исследова0 ний роль полиморфизма АДГ3 как фактора риска развития АБП ста0 вится под сомнение.
Генетические факторы, пред0 располагающие к возникновению АБП, также изучались на 158 ли0 цах, злоупотреблявших алкоголем [18]. Суточная доза для мужчин со0 ставила более 120 г алкоголя, для женщин – свыше 60 г. Проводились следующие генетические исследо0 вания: анализ 9 полиморфных ре0 гионов, картированных в экзонах III
иIX гена АДГ2, в экзоне VIII гена АДГ3, в интроне VI и области про0 мотера гена CYP2E1, а также в об0 ласти промотера гена туморне0 кротизирующего фактора α (TNF0α).
Узлоупотреблявших алкоголем лиц как с АБП, так и без нее выяв0 лены достоверные различия в рас0 пределении аллелей генов CYP2E1
иАДГ3. У жителей одного из двух включенных в исследование горо0 дов аллель С2 в промотерном ре0 гионе гена CYP2E1 встречался с частотой 0,06 у здоровых, злоупо0 треблявших алкоголем лиц, с час0 тотой 0,19 – у злоупотреблявших алкоголем и имевших АБП лиц (р=0,012) и с частотой 0,33 – у зло0 употреблявших алкоголем с цир0 розом печени (р=0,033). У жителей второго города была выявлена чет0 кая ассоциация между развитием АБП и гомозиготным наследовани0 ем аллеля АДГ3*2 гена АДГ3, его распространение среди злоупо0 треблявших алкоголем лиц с АБП составило 0,31, а среди здоровых, злоупотреблявших алкоголем, – 0,07 (р=0,004).
По результатам данного иссле0 дования гетерозиготность по алле0 лю С2 гена CYP2E1 и гомозигот0 ность по аллелю АДГ3*2 гена АДГ3 представляют собой независимые факторы риска развития АБП сре0 ди злоупотребляющих алкоголем лиц. Отсутствие этих аллелей сни0
жает риск развития АБП среди пьющих «опасные» дозы алкоголя в 3,2 и 4,3 раза соответственно.
Ассоциация между полимор0 физмом RELF гена Р450 2Е1 и АБП изучалась рядом авторов. Полу0 ченные результаты противоречивы.
Y. Maezawa и соавт. [16] ис0 следовали 82 японцев, страдав0 ших алкоголизмом (все мужчины). RELF человеческого гена Р450 2Е1 был выделен из лимфоцитов мето0 дом полимеразной цепной реак0 ции при воздействии рестриктиру0 ющих эндонуклеаз на последова0 тельности Rsa I (интрон 3) и Pst I. У 20 пациентов выявлена жировая дистрофия печени, у 60 – призна0 ки фиброза или цирроза. Тип А гена Р450 2Е1 (с1/с1) достоверно превалировал у пациентов с фиброзом и циррозом печени (р<0,05). Исследователи пришли к выводу о том, что восприимчи0 вость пациентов к тяжелому по0 вреждению печени может ассоци0 ироваться с Rsa I и Pst I полимор0 физмом гена Р450 2Е1. Эта точка зрения находит подтверждение и в других работах [11, 27].
Американские ученые Y.J. Wan и соавт. [30] также связывают раз0 витие АБП с Rsa I полиморфизмом гена Р450 2Е1 у американцев мек0 сиканского происхождения. С дру0 гой стороны, есть данные [21] о том, что генотип CYP2E1 не связан
сразвитием АБП в отличие от ге0 нотипа АЛДГ2, что также проде0 монстрировано на группе пациен0 тов японской национальности.
Результаты исследования поли0 морфизма в регуляторном регионе гена CYP2E1 на лицах белой расы также не выявили связи с развити0 ем АБП [22].
N.A. Wong и соавт. [31] изуча0 ли связь между Rsa I, Dra I, Tag I, генетическим полиморфизмом CYP2E1 и восприимчивостью к ал0 когольному повреждению печени или гепатоцеллюлярной карцино0 мой (ГЦК). Исследовали образцы ДНК, полученные от 61 пациента
сАБП, 41 пациента с ГЦК и 375 здоровых лиц группы контроля. Ме0 таанализ представлен по резуль0
татам предшествующих исследо0 ваний Rsa I полиморфизма и риска развития АБП.
В итоге работы не выявлено ас0 социации между полиморфизмом Rsa I, Dra I, Tag I и ГЦК. Распреде0 ление Rsa I и Dra I аллелей было сходным среди лиц с АБП и группы контроля. Какой0либо достовер0 ной связи между полиморфизмом Rsa I и Dra I и риском развития АБП не выявлено. Следует обратить внимание на то, что аллель Tag I достоверно реже встречался у лиц с АБП, чем в группе контроля. На основании этого высказано пред0 положение о вероятности его про0 тективной роли.
Генетическую предрасполо0 женность к АБП В России изучали П.П. Огурцов и соавт. [1, 19–21]. Предметом исследования стала частота аллеля АДГ2*2 в москов0 ской городской популяции и его корреляция с алкогольной зави0 симостью без признаков повреж0 дения печени и с развитием алко0 гольного цирроза печени. В ис0 следование были включены 123 москвича, из них группу контроля составили 50 здоровых доноров. У 36 человек имелся алкогольный цирроз печени, 37 страдали алко0 гольной зависимостью без при0 знаков повреждения печени. Час0 тота аллеля АДГ 2*2 для москов0 ской популяции составила 41%. Данная частота занимает сред0 нее положение между распрост0 раненностью этого аллеля у азиа0 тов и жителей Центральной и За0 падной Европы.
Существует негативная корре0 ляция между АДГ 2*2 и злоупотреб0 лением алкоголя как для больных с алкогольной зависимостью без при0 знаков повреждения печени, так и для больных с алкогольным цир0 розом печени. Четкой связи между риском развития цирроза печени и аллелем АДГ 2*2 не выявлено.
Заключение
Доказательство генетиче0 ской предрасположенности к раз0 витию тяжелых форм АБП позволи0
Гастроэнтерология, базирующаяся на доказательствах
9

Гастроэнтерология, базирующаяся на доказательствах
Клинические перспективы гастроэнтерологии, гепатологии ■ 5, 2004
ло бы прогнозировать риск разви0 тия этого заболевания у отдельных
Список литературы
1.Огурцов П.П. Вредные последствия потребления алкоголя в России: роль гене0 тического полиморфизма АДГ2 // Алко0 гольная политика России и Норвегии. – М.: Радуга, 2002. – С. 50–57.
2.Щипков В.П., Кривошеева Г.Н. Об0 щая и медицинская генетика. – М.: Изд. центр «Академия», 2003. – 256 с.
3.Carr L.G., Hartleroad J.Y., Liang Y. et al. Polymorphism at the P450 IIE1 locus is not assotiated with alcoholic liver disease in Caucasian men // Alcohol. Clin. Exp. Res. –1995. – Vol. 19, N 1. – P. 182–184.
4.Chao Y.0C., Liou S.0R., Chung Y.0Y. et al. Polymorphism of alcohol and aldehyde de0 hydrogenase genes and alcoholic cirrhosis in Chinese patients // Hepatology. – 1994. – Vol. 19. – P. 360–366.
5.Couzigou P., Fleury B., Groppi A. et al. Role of alcohol dehydrogenase polymorphism in ethanol metabolism and alcohol0related diseases // Adv. Exp. Med. Biol. – 1991. – Vol. 284. – P. 263–270.
6.Day C.P., James O.F.W., Bassendi0 ne M.F. et al. Alcohol dehydrogenase poly0 morphism and predisposition to alcoholic cirrhosis (Letter) // Hepatology. – 1993. – Vol. 18. – P. 230–232.
7.Day C.P., Bashir R., James O.F.W. et al. Investigation of the role of polymorphisms at the alcohol and aldehyde dehydrogenase loci in genetic predisposition to alcohol0related end0organ damage // Hepatology. – 1991.
–Vol. 13. – P. 798–801.
8.Ethanol and the Liver Mechanisms and Management / Ed. by D.I.N. Sherman, V. Pre0 edy, R.R. Watson. – N.Y.: Taylor & Francis, 2002. – 689 p.
9.Gilder F.J., Hodgkinson S., Murray R.M.
ADH and ALDH genotype profiles in Caucasians with alcohol0related problems and controls // Addiction. – 1993. – Vol. 88.
–P. 383–388.
10.Hrubec Z., Omenn G.S. Evidence of genetic predisposition to alcoholic cirrhosis and psychosis: twin concordances for alco0 holism and its biological endpoints by zygosi0 ty among male veterans // Alcohol. Clin. Exr. Res. – 1981. – Vol. 5. – P. 207–215.
11.Kato S., Onda M., Matsukura N. et al. Cytochrome P450 2E1 (CYP2E1) genetic poly0 morphism in a case0control study of gastric cancer and liver disease // Pharmacoge0 netics. – 1995. – Vol. 5, spec. N. – P. S141–S144.
12.Koivisto T., Mishin V.M., Mak K.M.
лиц. Однако результаты исследо0 |
вы, а сама проблема требует даль0 |
ваний в этой области противоречи0 |
нейшего изучения. |
et al. Induction of cytochrome P450 2E1 by ethanol in ret Kupffer cells // Alcohol. Clin. Exp. Res. – 1996. – Vol. 20. – P. 207–212.
13.Kopun M., Propping P. The kinetics of ethanol absorption and elimination in twins and supplementary repetitive experiments in singleton subjects // Europ. J. Clin. Phar0 macol. – 1977. – Vol. 11. – P. 337–344.
14.Kuntz E., Kuntz H.0D. Hepatology Principles and Practice. Berlin; Heidelberg: Springer0Verlag, 2002. – P. 52–54.
15.Lieber C.S. Alcoholic liver disease: new insight in pathogenesis lead to new treat0 ments // J. Hepatology. – 2000. – Vol. 32, suppl 1. – P. 113–128.
16.Maezawa Y., Yamauchi M., Toda G.
Assotiation between restriction fragment length polymorphism of the human cytochrome P450 IIE1 gene and susceptibility to alcoholic liver cirrhosis // Amer. J. Gastroenterol. – 1994. – Vol. 89, N 4. – P. 561–565.
17.McCaver D.G., Thomasson H.R., Martier S.S. et al. Alcohol dehydrogenase02*3 allele protects against alcohol0related birth
defects among African Americans // J. Pharmacol. Exp. Ther. – 1997. – Vol. 283. –
P.1095–1101.
18.Monzoni A., Masutti F., Saccoccio G. et al. Genetic determinants of ethanol0 induced liver damage // Mol. Med. – 2001. – Vol. 7, N 4. – P. 255–262.
19.Ogurtsov P.P., Garmash I.V., Miandina G.I. et al. Alcohol Dehydrogenase ADH 201 and ADG 202 allelic isoforms in Russian population correlate with type of alco0 holic disease // Addiction. Biology. – 2001. – Vol. 6. – P. 377–383.
20.Ogurtsov P.P., Nuzny V.P., Gar0 mash I.V. et al. Mortality in Russia // Lancet. – 2001. – Vol. 358, N 9282. – Vol. 669–670.
21.Okamoto K., Murawaki Y., Yuasa I. et al. Effect of ALDH2 and CYP2E1 gene poly0 morphisms on drinking behavior and alcoholic liver disease in Japanese male workers // Alcohol. Clin. Exp. Res. – 2001. – Vol. 25, N 6, suppl. – P. 19S–23S.
22.Plee0Gautier E., Foresto F., Ferrara R. et al. Genetic repeat polymorphisms in the regulating region of CYP2E1: frequency and relationship with enzymatic activity in alco0 holics // Alcohol. Clin. Exp. Res. – 2001. – Vol. 25, N 6. – P. 800–804.
23.Poupon R.E., Nalpas B., Coutelle C. et al. Polymorphism of alcohol dehydroge0 nase, alcohol and aldehyde dehydrogenase activities: implication in alcoholic cirrhosis in
white patients // Hepatology. – 1992. – Vol. 15. – P. 1017–1022.
24.Raucy J.L., Schultz E.D., Kearins M.C. et al. CYP2E1 expression in human lympho0 cytes from various ethnic populations // Alcohol. Clin. Exp. Res. – 1999. – Vol. 23, N 12. – P. 1868–1874.
25.Reed R., Page W.F., Viken R.J., Christian J.C. Genetic predisposition to organ specific endpoins of alcoholism // Alcohol. Clin. Exp. – 1996. – Vol. 20. – P. 1528–1533.
26.Sherman D.I., Ward R.J., Yoshida A., Peters T.J. Alcohol and acetaldehyde de0 hydrogenase gene polymorphism and alco0 holism // EXS. – 1994. – Vol. 71. – P. 291–300.
27.Tanaka F., Shiratori Y., Yokosuka O. et al. Polymorphism of alcohol0metabolizing genes affects drinking behavior and alcoholic liver disease in Japanese men // Alcohol. Clin. Exp. Res. – 1997. – Vol. 21. – P. 596–601.
28.Tanaka F., Shiratori Y.,Yokosuka O. et al. High incidence of ADH2*1/ALDG2*1 genes among Japanese alcohol depen0 dents and patients with alcoholic liver di0 sease // Hepatology. – 1996. – Vol. 23.
–P. 234–239.
29.Thomasson H.R., Beard J., Li T.0K.
ADH gene polymorphisms are determinants of alcohol pharmacokinetics // Alcohol. Clin. Exp. Res. – 1995. – Vol. 19. – P. 1494–1499.
30.Wan Y.J., Poland R.E., Lin K.M.
Genetic polymorphism of CYP2E1, ADH2 and ALDH2 in Mexican // Amer. Genet. Test. – 1998. – Vol. 2, N 1. – P. 79–83.
31.Wong N.A., Rae F., Simpson K.J. et al. Genetic polymorphisms of cytochrome P450 2E1 and susceptibility to alcoholic liver dise0 ase and hepatocellular carcinoma in a white population: a study and literature review, including meta0analysis // Mol. Pathol. – 2000. – Vol. 53, N 2. – P. 88–93.
32.Yamauchi M., Maezawa Y., Toda G. et al. Association of a restriction fragment length polymorphism in the alcohol dehydro0 genase02 gene with Japanese alcoholic liver cirrhosis // J. Hepatol. – 1995. – Vol. 23. – P. 519–523.
33.Yano H., Tsutsomi M., Fukura M. et al. Study of cytochrome P450 2E1 mRNA level of mononuclear cells in patients with alcoholic liver disease // Clin. Exp. Res. – 2001. – Vol. 25, N 6, suppl. – P. 2S–6S.
34.Zhang S., Lui S., Liu H., Zhu W.
Genotyping cytochrome P450 IIE1 in alco0 holic liver disease and its significance // Zhonghua Gan Zang Bing Za Zhi. – 2000. – Vol. 8, N 6. – P. 338–339.
10