

ХИМИЯ И ОБМЕН ЛИПИДОВ
Липиды представляют собой обширную группу соединений, различающихся по своей химической структуре и функциям.
Можно сказать, что липиды представляют собой группу веществ, которые характеризуются следующими признаками: нерастворимостью в воде; растворимостью в неполярных растворителях (эфир, хлороформ или бензол); содержанием высших алкильных радикалов; распространенностью в живых организмах.
Под это определение попадает большое количество веществ, в том числе такие, которые обычно причисляют к другим классам соединений: например, жирорастворимые витамины и их производные, каротиноиды, высшие углеводороды и спирты. Включение всех этих веществ в число липидов в известной степени оправдано, потому что в живых организмах они находятся вместе с липидами и вместе с ними экстрагируются неполярными растворителями. С другой стороны, имеются представители липидов, которые довольно хорошо растворяются в воде (например, лизолецитины). Термин «липиды» является более общим, чем термин «липоиды», который объединяет группу жироподобных веществ, таких, как фосфолипиды, стерины,
сфинголипиды и др.
БИОЛОГИЧЕСКАЯ РОЛЬ ЛИПИДОВ
Липиды играют важнейшую роль в процессах жизнедеятельности. Будучи одним из основных компонентов биологических мембран, липиды влияют на их проницаемость, участвуют в передаче нервного импульса, создании межклеточных контактов. Жир служит в организме весьма эффективным источником энергии либо при непосредственном использовании, либо потенциально – в форме запасов жировой ткани. В натуральных пищевых жирах содержатся жирорастворимые витамины и «незаменимые» жирные кислоты. Важная функция липидов – создание термоизоляционных покровов у животных и растений, защита органов и тканей от
механических воздействий.
КЛАССИФИКАЦИЯ ЛИПИДОВ
Наибольшее распространение получила классификация, основанная на структурных особенностях липидов. По этой классификации различают следующие основные классы липидов.
A. Простые липиды: сложные эфиры жирных кислот с различными спиртами.
1.Глицериды (ацилглицерины, или ацилглицеролы – по международной номенклатуре) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот.
2.Воска: сложные эфиры высших жирных кислот и одноатомных или двухатомных спиртов.
Б. Сложные липиды: сложные эфиры жирных кислот со спиртами, дополнительно содержащие и другие группы.
1.Фосфолипиды: липиды, содержащие, помимо жирных кислот и спирта, остаток фосфорной кислоты. В их состав часто входят азотистые основания и другие компоненты:
а) глицерофосфолипиды (в роли спирта выступает глицерол); б) сфинголипиды (в роли спирта – сфингозин).
2.Гликолипиды (гликосфинголипиды).
3.Стероиды.
4.Другие сложные липиды: сульфолипиды, аминолипиды. К этому классу можно отнести и липопротеины.
B. Предшественники и производные липидов: жирные кислоты, глицерол, стеролы и прочие спирты (помимо глицерола и стеролов), альдегиды жирных кислот, углеводороды, жирорастворимые витамины и гормоны.

ЖИРНЫЕ КИСЛОТЫ
Жирные кислоты – алифатические карбоновые кислоты – в организме могут находиться в свободном состоянии (следовые количества в клетках и тканях) либо выполнять роль строительных блоков для большинства классов липидов *.
В природе обнаружено свыше 200 жирных кислот, однако в тканях человека и животных в составе простых и сложных липидов найдено около 70 жирных кислот, причем более половины из них в следовых количествах. Практически значительное распространение имеют немногим более 20 жирных кислот. Все они содержат четное число углеродных атомов, главным образом от 12 до 24. Среди них преобладают кислоты, имеющие С16 и С18 (пальмитиновая, стеариновая, олеиновая и линолевая). Нумерацию углеродных атомов в жирно-кислотной цепи начинают с атома углерода карбоксильной группы. Примерно 3/4 всех жирных кислот являются непредельными (ненасыщенными), т.е. содержат двойные связи. Ненасыщенные жирные кислоты человека и животных, участвующие в построении липидов, обычно содержат двойную связь между (9-м и 10-м атомами углеводородов); дополнительные двойные связи чаще бывают на участке между 11-м атомом углерода и метильным концом цепи. Своеобразие двойных связей природных ненасыщенных жирных кислот заключается в том, что они всегда отделены двумя простыми связями, т.е. между ними всегда имеется хотя бы одна метиленовая группа (—СН=СН—СН2—СН=СН—). Подобные двойные связи обозначают как «изолированные».
Систематическое название жирной кислоты чаще всего образуется путем добавления к названию углеводорода окончания -овая. Насыщенные кислоты при этом имеют окончание -ановая (например, октановая кислота – систематическое название, каприловая кислота – тривиальное название), а ненасыщенные кислоты – -еновая (например, октадеценовая кислота
– систематическое название, олеиновая кислота – тривиальное название) .
В соответствии с систематической номенклатурой количество и положение двойных связей в ненасыщенных жирных кислотах часто обозначают с помощью цифровых символов: например, олеиновую кислоту как 18:1;9, линолевую кислоту как 18:2;9,12, где первая цифра – число углеродных атомов, вторая – число двойных связей, а следующие цифры – номера ближайших к карбоксилу углеродных атомов, вовлеченных в образование двойной связи.
В специальной литературе жирные кислоты часто изображают в виде зигзагообразной вытянутой линии, отражающей жесткость валентного угла атомов углерода в 111° для насыщенной и в 123° – для двойной связи. Такая конформация является условной и справедлива только для случая, когда жирная кислота находится в кристаллическом состоянии.
*В природе значительно чаще встречаются длинноцепочечные жирные кислоты с числом углеродных атомов больше двенадцати, часто их называют - «высшие жирные кислоты».

В растворах жирно-кислотная цепь может образовывать бесчисленное количество конформаций вплоть до клубка, в котором имеются и линейные участки различной длины в зависимости от числа двойных связей. Клубки могут слипаться между собой, образуя так называемые мицеллы. В последних отрицательно заряженные карбоксильные группы жирных кислот обращены к водной фазе, а неполярные углеводородные цепи спрятаны внутри мицеллярной структуры. Такие мицеллы имеют суммарный отрицательный заряд и в растворе остаются суспендированными благодаря взаимному отталкиванию.
Известно также, что при наличии двойной связи в жирнокислотной цепи вращение углеродных атомов относительно друг друга ограничено. Это обеспечивает существование ненасыщенных жирных кислот в виде геометрических изомеров, причем природные ненасыщенные жирные кислоты имеют цис-конфигурацию и крайне редко транс-конфигурации. Считают, что жирной кислоте с несколькими двойными связями цисконфигурация придает углеводородной цепи изогнутый и укороченный вид. По этой причине молекулы этих кислот занимают больший объем, а при образовании кристаллов упаковываются не так плотно, как транс-изомеры. Вследствие этого цис-изомеры имеют более низкую температуру плавления (олеиновая кислота, например, при комнатной температуре находится в жидком состоянии, тогда как элаидиновая – в кристаллическом). Цисконфигурация делает ненасыщенную кислоту менее стабильной и более подверженной катаболизму.
ГЛИЦЕРИДЫ (АЦИЛГЛИЦЕРОЛЫ)
Глицериды (ацилглицерины, или ацилглицеролы*) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот. Если жирными кислотами этерифицированы все три гидроксильные группы глицерина (ацильные радикалы R1 , R2 и R3 могут быть одинаковы или различны), то такое соединение называют триглицеридом (триацилглицерол), если две – диглицеридом (диацилглицерол) и если этерифицирована одна
группа – моноглицеридом (моноацилглицерол):
Наиболее распространенными являются триглицериды, часто называемые нейтральными жирами или просто жирами. Нейтральные жиры находятся в организме либо в форме протоплазматического жира, являющегося структурным компонентом клеток, либо в форме запасного, резервного, жира. Роль этих двух форм жира в организме неодинакова. Протоплазматический жир имеет постоянный химический состав и содержится в тканях в определенном количестве, не изменяющемся даже при патологическом ожирении, в то время как количество резервного жира подвергается большим колебаниям.
Жирные кислоты в триглицеридах могут быть насыщенными и ненасыщенными. Из жирных кислот чаще встречаются пальмитиновая, стеариновая и олеиновые кислоты. Если все три кислотных радикала принадлежат одной и той же жирной кислоте, то такие триглицериды называют простыми (например, трипальмитин, тристеарин, триолеин и т.д.), если разным жирным кислотам, то смешанными.

Названия смешанных триглицеридов образуются в зависимости от входящих в их состав жирных кислот, при этом цифры 1, 2 и 3 указывают на связь остатка жирной кислоты с соответствующей спиртовой группой в молекуле глицерина (например, 1-олео-2- пальмитостеарин). Положение крайних атомов в молекуле глицерина на первый взгляд равнозначно, тем не менее их обозначают сверху вниз – 1 и 3. Это объясняется тем, что в структуре триглицерида при пространственном ее рассмотрении крайние «глицериновые» атомы углерода становятся уже не равнозначными, если гидроксилы 1 и 3 ацилированы разными жирными кислотами. При необходимости применяют также систему стереохимической нумерации (обозначают sn – stereochemical numbering): например, 1,2-дистеарил-3-пальмитил- sn-глицерол:
По этой системе, если в проекции Фишера гидроксильная группа при 2-м углеродном атоме глицерина располагается слева, атому углерода, находящемуся над ним, присваивается номер 1, а расположенному под ним – номер 3.
Действительно, углероды 1 и 3 глицерола, учитывая их пространственное расположение, неидентичны. Особенно четко это видно на примере молекулы триглицерида. Ферменты это различают и всегда специфичны только к одному из трех углеродов глицерина. Так, глицеролкиназа фосфорилирует глицерин в положении sn-3, в результате чего образуется глицерол-3-фосфат, но не глицерол-1-фосфат.
Жирные кислоты, входящие в состав триглицеридов, практически определяют их физикохимические свойства. Так, температура плавления триглицеридов повышается с увеличением числа и длины остатков насыщенных жирных кислот. Напротив, чем выше содержание ненасыщенных жирных кислот, или кислот с короткой цепью, тем ниже точка плавления.
Животные жиры (сало) обычно содержат значительное количество насыщенных жирных кислот (пальмитиновой, стеариновой и др.). благодаря чему при комнатной температуре они твердые. Жиры, в состав которых входит много ненасыщенных кислот, при обычной температуре жидкие и называются маслами. Так, в конопляном масле 95% всех жирных кислот приходится на долю олеиновой, линолевой и линоленовой кислот и только 5% – на долю стеариновой и пальмитиновой кислот. В жире человека, плавящемся при температуре 15°С (при температуре тела он жидкий), содержится 70% олеиновой кислоты.
Глицериды способны вступать во все химические реакции, свойственные сложным эфирам. Наибольшее значение имеет реакция омыления, в результате которой из триглицеридов образуются глицерол и жирные кислоты. Омыление жира может происходить как при ферментативном гидролизе, так и при действии кислот или щелочей.
Жиры в качестве примесей содержат небольшое количество свободных жирных кислот и незначительное количество неомыляемых веществ.
ВОСКА
Воска – сложные эфиры высших жирных кислот и высших одноатомных или двухатомных спиртов с числом углеродных атомов от 16 до 22. Общие их формулы можно представить так:
Воска могут входить в состав жира, покрывающего кожу, шерсть, перья. У растений 80% от всех липидов, образующих пленку на поверхности листьев и плодов, составляют воска.

Известно также, что воска являются нормальными метаболитами некоторых микроорганизмов. Природные воска (например, пчелиный воск, спермацет, ланолин) обычно содержат, кроме указанных сложных эфиров, некоторое количество свободных жирных кислот, спиртов и
углеводородов с числом углеродных атомов 21–35.
ФОСФОЛИПИДЫ
Фосфолипиды представляют собой сложные эфиры многоатомных спиртов глицерина или сфингозина с высшими жирными кислотами и фосфорной кислотой. В состав фосфолипидов входят также азотсодержащие соединения: холин, этаноламин или серин. В зависимости от того, какой многоатомный спирт участвует в образовании фосфолипида (глицерин или сфингозин), последние делят на 2 группы: глицерофосфолипиды и сфингофосфолипиды. В глицерофосфолипидах либо холин, либо этаноламин или серин соединены эфирной связью с остатком фосфорной кислоты; в составе сфинголипидов обнаружен только холин. Наиболее распространенными в тканях животных являются глицерофосфолипиды.
Глицерофосфолипиды
Являются производными фосфатидной кислоты. В их состав входят глицерин, жирные кислоты, фосфорная кислота и обычно азотсодержащие соединения. Общая формула:
Вэтих формулах R1 и R2 – радикалы высших жирных кислот, a R3 – чаще радикал азотистого соединения. Для всех глицерофосфолипидов характерно, что одна часть их молекул (радикалы R1 и R2 ) обнаруживает резко выраженную гидрофобность, тогда как другая часть гидрофильна благодаря отрицательному заряду фосфорной кислоты и положительному заряду радикала R3 .
Из всех липидов глицерофосфолипиды обладают наиболее выраженными полярными свойствами. При помещении глицерофосфолипидов в воду в истинный раствор переходит лишь небольшая их часть, основная же масса липидов находится в виде мицелл.
Взависимости от характера азотистого основания, присоединенного к фосфорной кислоте, Глицерофосфолипиды подразделяют на фосфатидилхолины (лецитины), фосфатидилэтаноламины (кефалины) и фосфатидилсерины. В состав некоторых глицерофосфолипидов вместо азотсодержащих соединений входит не содержащий азота шестиуглеродный циклический спирт инозит, называемый также инозитолом. Эти липиды называются фосфатидилинозитолами.
1.Фосфатидилхолины (лецитины). В отличие от триглицеридов в молекуле фосфатидилхолина одна из трех гидроксильных групп глицерина связана не с жирной, а с фосфорной кислотой. Кроме того, фосфорная кислота в свою очередь соединена эфирной связью с азотистым основанием – холином [НО—СН2—СН2—N+ (CH3 )3 ]. Таким образом, в молекуле фосфатидилхолина соединены глицерин, высшие жирные кислоты, фосфорная кислота и холин:
2. Фосфатидилэтаноламины. Основное различие между фосфатидилхолинами и фосфатидилэтаноламинами – наличие в составе последних азотистого основания этаноламина
(HO—CH2—CH2—N+H3 ):

Из глицерофосфолипидов в организме животных и высших растений в наибольшем количестве встречаются фосфатидилхолины и фосфатидилэтаноламины. Эти 2 группы глицерофосфолипидов метаболически связаны друг с другом и являются главными липидными компонентами мембран клеток.
3. Фосфатидилсерины. В молекуле фосфатидилсерина азотистым соединением служит остаток аминокислоты серина
Фосфатидилсерины распространены гораздо менее широко, чем фосфатидилхолины и фосфоэтаноламины, их значение: участвуют в синтезе фосфатидилэтаноламинов.
4. Фосфатидилинозитолы. Эти липиды относятся к группе производных фосфатидной кислоты, но не содержат азота. Радикалом (R3) в этом подклассе глицерофосфолипидов является шестиуглеродный циклический спирт инозитол:
Фосфатидилинозитолы довольно широко распространены в природе. Они обнаружены у животных, растений и микроорганизмов. В животном организме найдены в мозге, печени и легких.
5. Плазмалогены. От рассмотренных глицеролипидов плазмалогены отличаются тем, что вместо одного остатка высшей жирной кислоты содержат остаток α,β-ненасыщенного спирта, который образует простую связь (в отличие от сложноэфирной связи, образуемой остатком жирной кислоты) с гидроксильной группой глицерина в положении С-1:
Основными подклассами плазмалогенов являются фосфатидальхолины, фосфатидальэтаноламины и фосфатидальсерины. В разбавленных кислотах плазмалогены гидролизуются с образованием альдегида соответствующего α,β-ненасыщенного спирта, т.е. при кислотном гидролизе плазмалогенов образуются «жирные» альдегиды, называемые плазмалями, что и легло в основу термина «плазмалоген».
6. Кардиолипин. Впервые выделен из сердечной мышцы. Это соединение, в котором 2 молекулы фосфатидной кислоты связаны с помощью одной молекулы глицерина. В отличие от остальных глицерофосфолипидов кардиолипин является как бы «двойным» глицерофосфолипидом. Кардиолипин локализован во внутренней мембране митохондрий. Функция его пока неясна, хотя известно, что в отличие от других фосфолипидов кардиолипин обладает иммунными свойствами
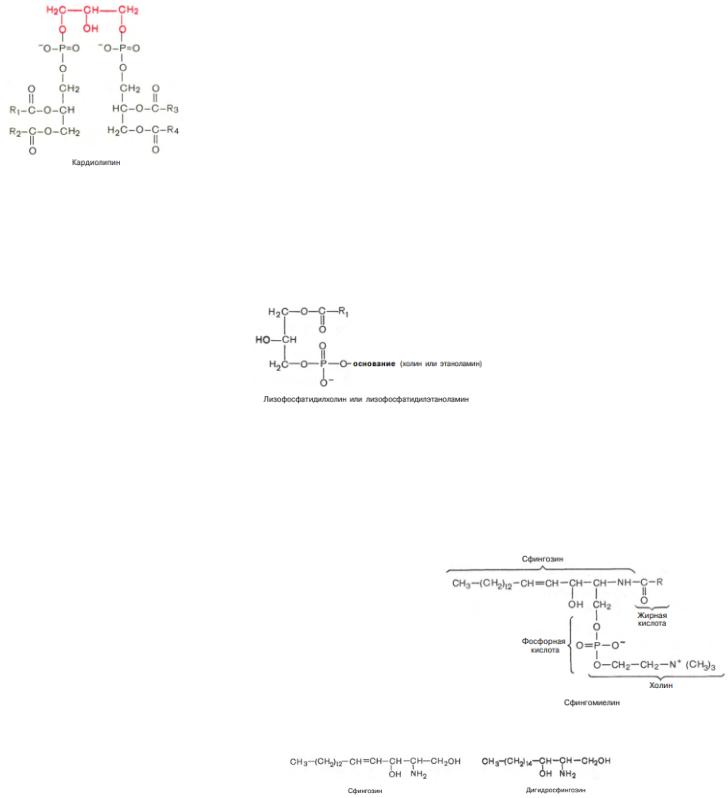
Необходимо отметить, что в природе встречается свободная фосфатидная кислота, но в относительно небольших количествах по сравнению с глицерофосфолипидами. Среди жирных кислот, входящих в состав глицерофосфолипидов, обнаружены как насыщенные, так и ненасыщенные (чаще стеариновая, пальмитиновая, олеиновая и линолевая).
Большинство фосфатидилхолинов и фосфатидилэтаноламинов содержат одну насыщенную высшую жирную кислоту в положении С-1 и одну ненасыщенную высшую жирную кислоту в положении С-2. Гидролиз фосфатидилхолинов и фосфатидилэтаноламинов при участии особых ферментов (эти ферменты относятся к фосфолипазам А2 ), содержащихся, например, в яде кобры, приводит к отщеплению ненасыщенной жирной кислоты и образованию лизофосфолипидов – лизофосфатидилхолинов, или лизофосфатидилэтаноламинов, оказывающих сильное гемолитическое действие:
Сфинголипиды (сфингофосфолипиды)
Сфингомиелины. Наиболее распространенные сфинголипиды. В основном они находятся в мембранах животных и растительных клеток. Особенно богата ими нервная ткань. Сфингомиелины обнаружены также в ткани почек, печени и других органов. При гидролизе сфингомиелины образуют одну молекулу жирной кислоты, одну молекулу двухатомного ненасыщенного спирта сфингозина, одну молекулу азотистого основания (чаще это холин) и одну молекулу фосфорной кислоты. Общая формула:
Молекула сфингомиелина содержит как бы полярную «головку», которая несет одновременно и положительный (остаток холина), и отрицательный (остаток фосфорной кислоты) заряды, и два неполярных «хвоста» (длинная алифатическая цепь сфингозина и ацильный радикал жирной кислоты). В некоторых сфингомиелинах, например выделенных из мозга и селезенки, вместо сфингозина найден спирт дигидросфингозин (восстановленный сфингозин):
ГЛИКОЛИПИДЫ (ГЛИКОСФИНГОЛИПИДЫ)
Гликолипиды широко представлены в тканях, особенно в нервной ткани, в частности в мозге. Главной формой гликолипидов в животных тканях являются гликосфинголипиды. Последние содержат церамид, состоящий из спирта сфингозина и остатка жирной кислоты, и один или несколько остатков сахаров. Простейшими гликосфинголипидами являются галактозилцерамиды и глюкозилцерамиды. Галактозилцерамиды * – главные сфинголипиды мозга и других нервных тканей, но в небольших количествах встречаются и во многих других тканях. В состав галактозилцерамидов входит гексоза (обычно это D-галактоза), которая связана эфирной связью с гидроксильной группой аминоспирта сфингозина. Кроме того, в составе галактозилцерамида имеется жирная кислота. Чаще всего это лигноцериновая, нервоновая или цереброновая кислота, т.е. жирные кислоты, имеющие 24 углеродных атома.
* Производные галактозилцерамидов нередко называют цереброзидами.
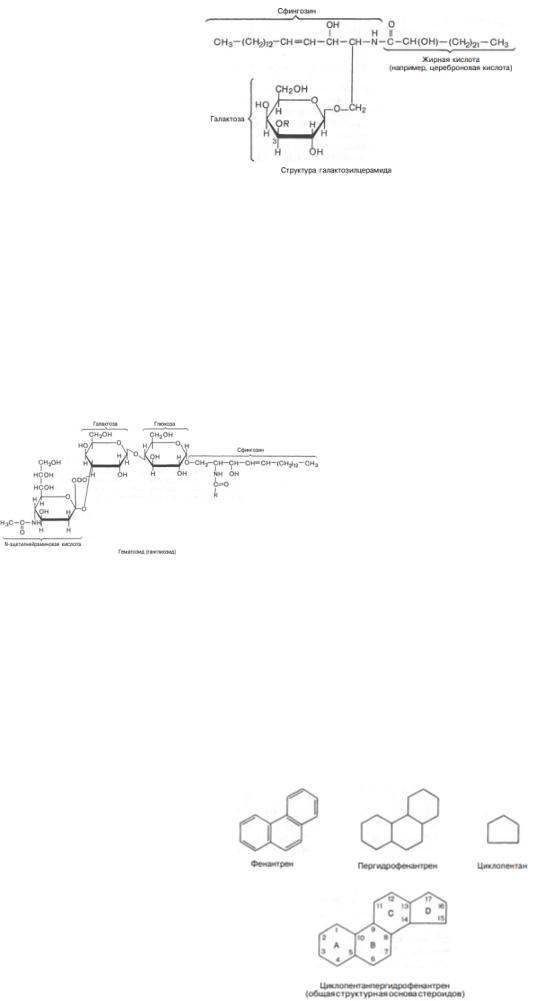
Существуют сульфогалактозилцерамиды, которые отличаются от галактозилцерамидов наличием остатка серной кислоты, присоединенного к третьему углеродному атому гексозы. В мозге млекопитающих сульфогалактозилцерамиды в основном находятся в белом веществе, при этом содержание их в мозге намного ниже, чем галактозилцерамидов.
Глюкозилцерамиды – простые гликосфинголипиды, представлены в тканях, отличных от нервной, причем главным образом глюкозилцерамидами. В небольших количествах они имеются в ткани мозга. В отличие от галактозилцерамидов у них вместо остатка галактозы имеется остаток глюкозы.
Более сложными гликосфинголипидами являются ганглиозиды, образующиеся из гликозилцерамидов. Ганглиозиды дополнительно содержат одну или несколько молекул сиаловой кислоты. В тканях человека доминирующей сиаловой кислотой является нейраминовая. Вместо остатка глюкозы они чаще содержат сложный олигосахарид. Ганглиозиды в больших количествах находятся в нервной ткани. Они выполняют рецепторные и другие функции. Одним из простейших ганглиозидов является гематозид, выделенный из стромы эритроцитов. Он содержит церамид (ацилсфингозин), одну молекулу глюкозы, одну молекулу N-ацетилнейраминовой кислоты
СТЕРОИДЫ
Все рассмотренные липиды принято называть омыляемыми, поскольку при их щелочном гидролизе образуются мыла. Однако имеются липиды, которые не гидролизуются с освобождением жирных кислот. К таким липидам относятся стероиды. Стероиды – широко распространенные в природе соединения. Они часто обнаруживаются в ассоциации с жирами. Их можно отделить от жира путем омыления* (они попадают в неомыляемую фракцию). Все стероиды в своей структуре имеют ядро, образованное гидрированным фенантреном (кольца А, В и С) и циклопентаном (кольцо D):
Омылением называется гидролиз жира щелочью. Продуктами омыления являются глицерин и щелочные соли жирных кислот, которые называют
мылами.
К стероидам относятся, например, гормоны коркового вещества надпочечников, желчные кислоты, витамины группы D, сердечные гликозиды и другие соединения. В организме человека важное место среди стероидов занимают стерины (стеролы), т.е. стероидные спирты. Главным представителем стеринов является холестерин (холестерол). Ввиду сложного строения и асимметрии молекулы стероиды имеют много потенциальных стереоизомеров. Каждое из шестиуглеродных колец (кольца А, В и С) стероидного ядра может принимать две различные пространственные конформации – конформацию «кресла» либо «лодки».
В природных стероидах, в том числе и в холестерине, все кольца в форме «кресла» (рис. 6.2), что является более устойчивой конформацией. В свою очередь по отношению друг к другу кольца могут находиться в цисили транс-положениях.
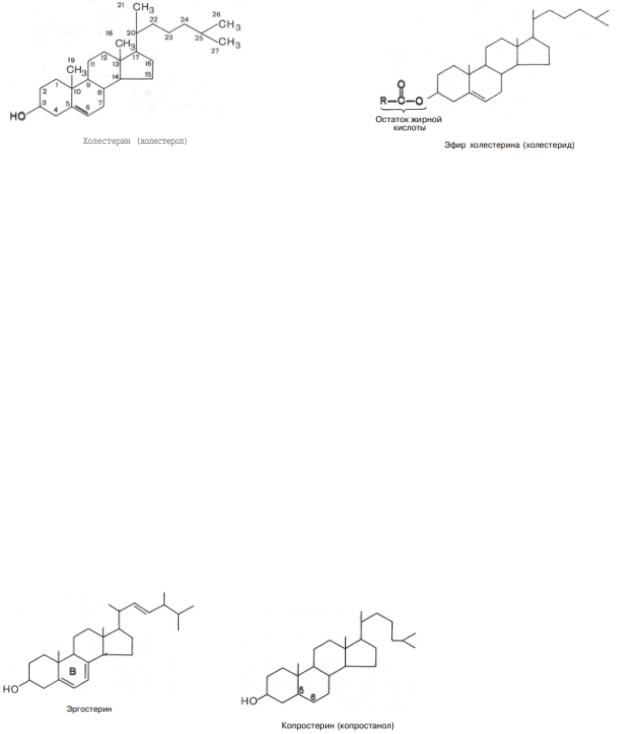
Холестерин. Среди стероидов выделяется группа соединений, получивших название стеринов (стеролов). Для стеринов характерно наличие гидроксильной группы в положении 3, а также боковой цепи в положении 17. У важнейшего представителя стеринов – холестерина – все кольца находятся в транс-положении; кроме того, он имеет двойную связь между 5-м и 6-м углеродными атомами. Следовательно, холестерин является ненасыщенным спиртом:
Кольцевая структура холестерина отличается значительной жесткостью, тогда как боковая цепь – относительной подвижностью. Итак, холестерин содержит спиртовую гидроксильную группу при С-3 и разветвленную алифатическую цепь из 8 атомов углерода при С-17. Химическое название холестерина 3-гидрокси-5,6-холестен. Гидроксильная группа при С3 может быть этерифицирована высшей жирной кислотой, при этом образуются эфиры холестерина (холестериды).
Каждая клетка в организме млекопитающих содержит холестерин. Находясь в составе мембран клеток, неэтерифицированный холестерин вместе с фосфолипидами и белками обеспечивает избирательную проницаемость клеточной мембраны и оказывает регулирующее влияние на состояние мембраны и на активность связанных с ней ферментов. В цитоплазме холестерин находится преимущественно в виде эфиров с жирными кислотами, образующих мелкие капли – так называемые вакуоли. В плазме крови как неэтерифицированный, так и этерифицированный холестерин транспортируется в составе липопротеинов. Холестерин – источник образования в организме млекопитающих желчных кислот, а также стероидных гормонов (половых и кортикоидных).
Холестерин, а точнее продукт его окисления – 7-дегидрохолестерин, под действием УФ-лучей в коже превращается в витамин D3 .
Холестерин находится в животных, но не в растительных жирах. В растениях и дрожжах содержатся близкие по структуре к холестерину соединения, в том числе эргостерин.
Эргостерин – предшественник витамина D. После воздействия на эргостерин УФ-лучами он приобретает свойство оказывать противорахитное действие (при раскрытии кольца В). Восстановление двойной связи в молекуле холестерина приводит к образованию копростерина (копростанола). Копростерин находится в составе фекалий и образуется в результате восстановления бактериями кишечной микрофлоры двойной связи в холестерине между атомами С5 и С6.
Указанные стерины в отличие от холестерина очень плохо всасываются в кишечнике и потому обнаруживаются в тканях человека в следовых количествах.