

БИОХИМИЯ, 2 СЕМИНАР (07.09.2020)
ОБЩАЯ ХАРАКТЕРИСТИКА ФЕРМЕНТОВ:
Ферменты - это биологические катализаторы. Все они по своей природе являются белками, некоторые содержат ещё и небелковую часть – кофактор.
СХОДСТВО И РАЗЛИЧИЕ В ДЕЙСТВИИ ФЕРМЕНТОВ И НЕОРГАНИЧЕСКИХ КАТАЛИЗАТОРОВ:
КЛАССИФИКАЦИЯ И НОМЕНКЛАТУРА ФЕРМЕНТОВ:
НОМЕНКЛАТУРЫ:
Субстрат : Химическое наименование акцептор - Типа рекции + -аза
Пример: L-аланин (субстрат) 2-оксоглутарат (ацептор) – аминотрансфераза (переносит аминогруппу с аланина на глутарат)
Рабочее название: сукцинатдегидрогеназа
КЛАССИФИКАЦИЯ:
1)Оксидоредуктазы: донор электронов : акцептор электронов – оксидоредуктаза
Катализируют окислительно-восстановительные реакции, лежащие в основе биологического окисления. Чаще всего ты будешь встречать их рабочие названия по окончаниям: дегидрогеназа, редуктаза, окидаза, монооксигеназ, пероксидаза и тд.
Коферменты: НАД, НАДФ, ФАД, ФМН, убихинон, глутатион.
ОН => С=О => ОВР => оксидоредуктаза.
Систематическое название: Алкоголь : НАД – оксидоредуктаза
Рабочее название: Алкогольдегидрогеназа
2)Трансферазы: донор группы : акцептор группы – трансфераза
Катализируют реакции переноса различных групп от одного субстрата (донор) к другому (акцептор), участвуют в реакции взаимопревращения различных веществ, обезвреживания природных и чужеродных соединений.
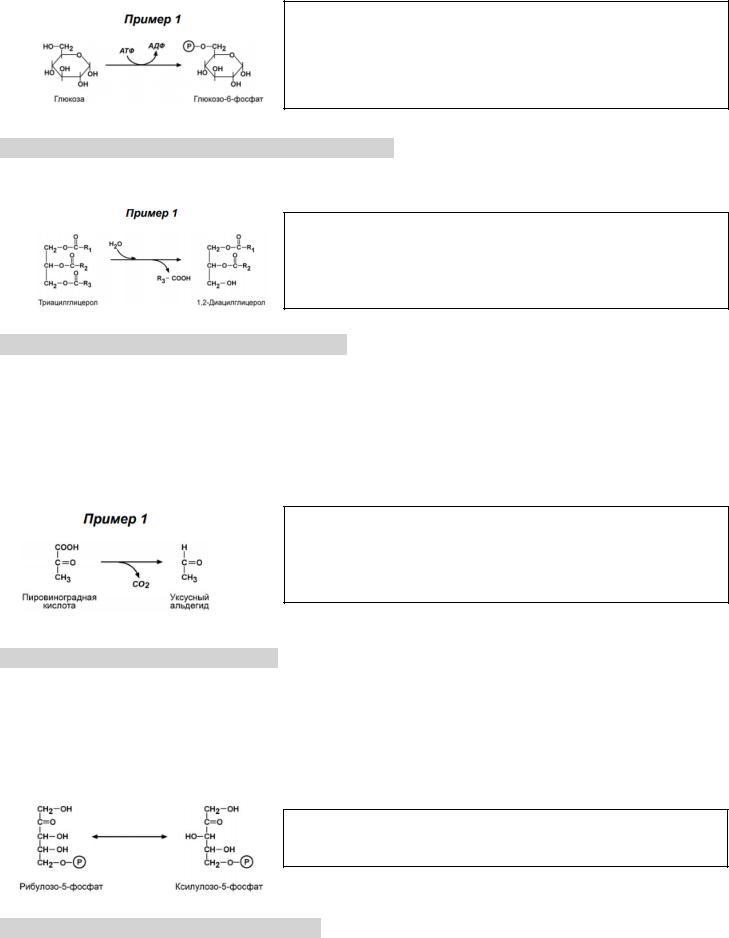
Коферменты: пиридоксальфосфат, коэнзим А, тетрагидрофолиевая кислота, метилкобаламин.
Глюкоза + Фосфатная группа (с АТФ) => Глюкоза-6-фосфат => трансфераза
Систематическое название: АТФ : Глюкоза – трансфераза
Рабочее название: Гексогеназа
3)Гидролазы: нет коферментов (ферменты простые)
Катализирует разрыв внутримолекулярных связей в субстрате путём присоединение воды.
Триацилглицерол => Гидролиз => 1,2-диацилглицерол => гидролаза
Систематическое название: Жир? => Липаза; ДНК? => ДНКаза и т.д.
4) Лиазы: фермент : отделяемая группа – лиаза
Катализируют разрыв углерод-углеродной связи, углерод-кислородной и углерод-азотной связи, и, в отличие от гидролаз, катализируют реакции отщепления различных групп уже негидролитическим путём. Часто реакции сопровождаются образованием двойной связи или присоединением групп к месту двойной связи.
Коферменты: пиридоксальфосфат, тиаминдифосфат, магний, кобальт
СООН от ПВК => СО2 => лиаза
Систематическое название: 2-оксокислота : карбокси – лиаза
Рабочее название: Пируватдекарбоксилаза
5) Изомеразы: субстрат – [] – реакция
Катализируют изомерные превращения в пределах одной молекулы.
Коферменты: пиридоксальфосфат, дезоксиаденозилкобаламин, глутатион, фосфаты моносахаридов. Рацемаза – стереоизомеризация; Эпимераза – эпимерия; Мутаза – цис- и трансизомеризация.
Рибулоза-5-фосфат => ксилулоза-5-фосфат => эпимераза
Систематическое название: Рибулоза-5-фосфат-эпимераза
6) Лигазы: субстрат 1 : субстрат 2 – лигаза
Катализируют присоединение друг к другу двух молекул при помощи энергии АТФ. АТФ обладает
макроэргическими связями, они высокоэнергетические, что позволяет лигазам с огромной скоростью образовывать между молекулами новые связи.

Коферменты: нуклеотидные (УТФ), биотиновые (витамин Н), фолиевые коферменты.
Глутамат + Аммиак => Глутамин => Лигазы
Систематическое название: Глутамат : Аммиак – Лигаза
Рабочее название: Глутаминсинтаза
ПОНЯТИЕ ЭНЕРГЕТИЧЕСКОГО БАРЬЕРА РЕАКЦИИ И ЭНЕРГИИ АКТИВАЦИИ:
Ферменты помогают опустить границу энергетического барьера для упрощения и ускорения реакции.
Так как повышение температуры и давления, а также облучение в условиях нашего организма невозможны, используются катализаторы.
Таким образом, катализаторы
снижают значение энергии активации
(энергия, необходимая для начала химической реакции).
Энергия активации – это количество энергии, которое необходимо для перевода всех молекул 1 моля вещества в активное состояние.
Энергетический барьер – это такое количество энергии, которое необходимо преодолеть молекулам, чтобы вступить в химическое взаимодействие. Величина энергии активации равняется величине энергетического барьера.
Простетическая группа – неотделяемая часть
Кофермент (кофактор) – отделяемая часть
o производные витаминов: НАД, ФАД, ФМН, TПФ o нуклеотиды
o порфирины (гем)
o металлы (кофакторы): K+, Fe2+, Cu2+, Co2+, Zn2+, Mn2+, Mg2+, Ca2+
ЭТАПЫ ФЕРМЕНТАТИВНОГО КАТАЛИЗА:
Всё очень просто – субстрат (S) присоединяется к ферменту (E – энзим). Образуется ферментсубстратный комплекс(E-S). Происходит биохимическая реакция, в одну или несколько стадий (E-X) . Уже измененный комплекс превращается в комплекс фермент-продукт (P), и продукт отделяется от фермента.
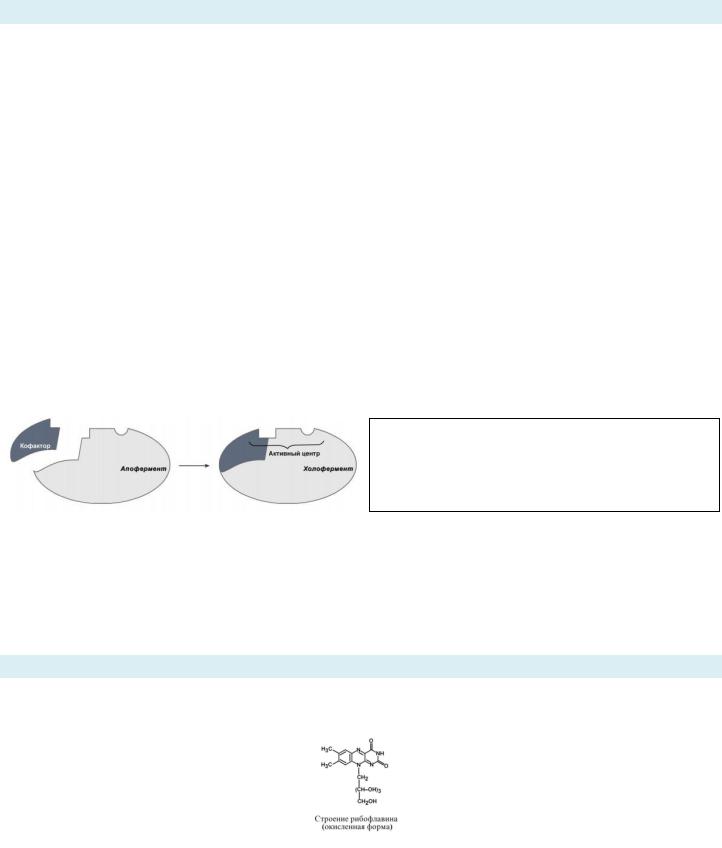
ХАРАКТЕРИСТИКА СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ ФЕРМЕНТОВ:
Простые ферменты = только белковая часть (АК) – апофермент.
Пример: пепсин, трипсин лизоцим и т.д.
Сложные ферменты (холоферменты) = белковая часть (апофермент) + небелковая часть (кофермент/кофактор/простетическая группа).
Пример: ферменты с РНК – рибозимы; с ФАД – суукцинатдегидрогеназа и т.д.
Активный центр - это определенный участок белковой молекулы, способный комплементарно связываться с субстратом и обеспечивающий его каталитическое превращение. Структура активного центра сформирована радикалами аминокислот, так же как и в случае активного центра любого белка. В активном центре фермента имеются аминокислотные остатки, функциональные группы которых обеспечивают комплементарное связывание субстрата (участок связывания), и аминокислотные остатки, функциональные группы которых осуществляют химическое превращение субстрата (каталитический участок).
Активный центр = Якорный участок (участок связвания) + Каталитический участок
Активный центр «активируется» при присоединение к апоферменту кофактора => полоценный холефермент с ЯУ и КУ => только такой тандем способен к катализу.
Аллостерический центр – он отделён от активного центра и обеспечивает регуляцию активности фермента. С аллостерическим центром связывается какая-либо молекула – ингибитор или активатор, модулятор, которые влияют на конфигурацию самого фермента - сродство активного центра к специфическому субстрату (лиганду) и регулирует его деятельность. Это может быть продукт данной или последующей реакций, субстрат реакции или другое вещество.
КОФЕРМЕНТНЫЕ ФОРМЫ ВИТАМИНОВ НА ПРИМЕРЕ В2 (ФМН и ФАД), РР (НАД+ и НАДФ+)
В2 (рибофлавин, витамин рост):
Кофермент оксидоредуктаз – обеспечивает перенос 2 атомов водорода в окислительновосстановительных реакциях.
Содержит в себе:
1.Дегидрогеназы энергетического обмена – пируватдегидрогеназа, альфакетоглутаратдегидрогеназа, сукцинатдегидрогеназа, ацил-КоА-дегидрогеназа, митохондриальная альфа-глицеролфосфатдегидрогеназа.
2.Оксидазы, окисляющие субстраты с участием молекулярного кислорода.
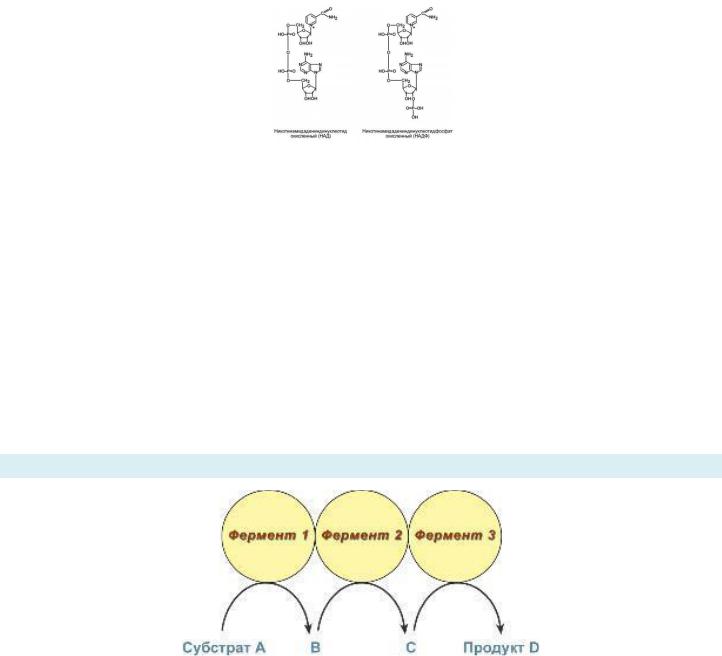
Состоит из флавина и спирта рибитола (нам это пока ненужно, но можешь блеснуть знаниями).
Чтобы он стал коферментом, он должен дополнительно содержать либо только фосфорную кислоту (тогда он превращается в коферментную форму - флавинмононуклеотид), либо фосфорную кислоту, дополнительно связанную с АМФ – флавинадениндинуклеотид (нуклеотид в составе рибофлавина и нуклеотид в составе АМФ, поэтом динуклеотид). В таком виде Витамин В2 может соединиться с активным центром фермента. В2 содержится в мясе, дрожжевых булочках и молочных продуктах.
РР (В3, ниацин, антипеллагрический):
Существует в виде никотиновой кислоты или никотинамида.
Его коферментные формы – всем нам давно известные ещё с первого курса НАД и НАДФ.
Биохимические функции: перенос гидрид-ионов Н в окислительно-восстановительных реакциях. То есть опять оксидоредуктазы.
Рассмотрим их названия:
oНАД: никотинамид (сам витамин В3) – аденин (азотистое основание) – динуклеотид (два нуклеотида в составе, у одного в качестве азотистого основания аденин, у другого –
никотинамид)
oНАДФ: никотинамид (витами В3) – аденин (азотистое основание) – динуклеотид – фосфат (рибоза в составе АМФ фосфорелированная)
ПОЯТИЕ О МУЛЬТИФЕРМЕНТАТИВНОМ КОМПЛЕКСЕ: СТРОЕНИЕ, ПРИНЦИПЫ САМОСБОРКИ, РОЛЬ
Ферменты лучше работают, когда соединены в комплексы. Они прочно связаны и осуществляют ряд последовательных реакций одного пути.
Работает это так: продукт реакции первого фермента с субстратом передаётся на следующий фермент и является только его субстратом.
Зачем эти комплексы нужны? Они значительно ускоряют ход реакции. Мультиферментные комплексы концентрируют каталитическую активность, повышают концентрацию интермедиатов, субстрат не
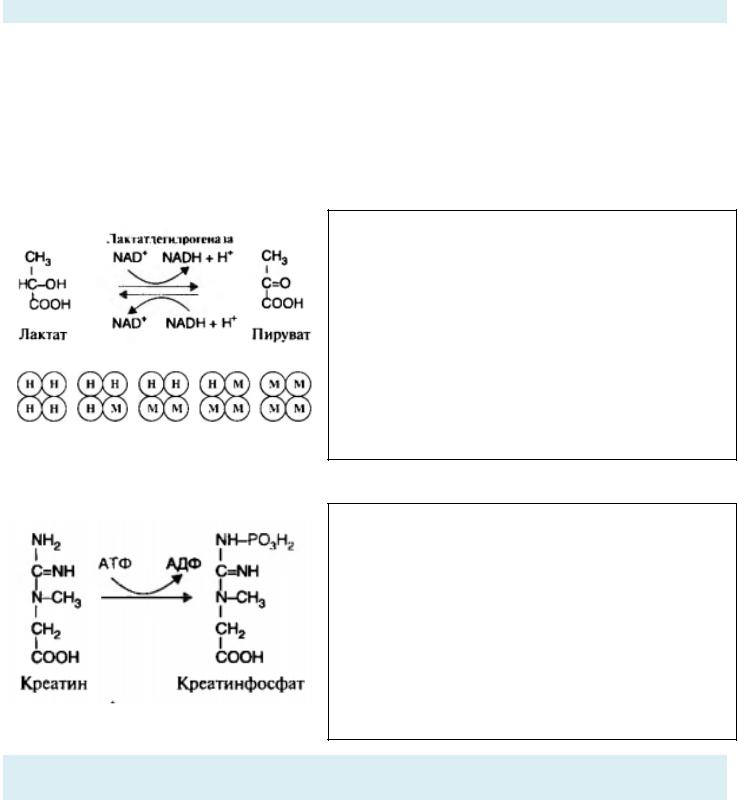
рассеивается из-за диффузного эффекта, а целиком переходит на следующий фермент, так сказать, безотходно.
Таких комплексов в нашем организме много: пируватдегидрогеназный комплекс (пируват-альфа- ацетил-S-КоА), альфа-кетоглутарадегидрогеназный комплекс (альфа-кетоглутарат-альфа-сукцинил-S- KoA), «синтаза жирных кислот» (синтез пальмитиновой кислоты).
ИЗОФЕРМЕНТЫ, ОСОБЕННОСТИ ИХ СТРОЕНИЯ НА ПРИМЕРЕ ЛАКТОДЕГИДРОГЕНАЗЫ И КРЕАТИНКИНАЗЫ:
Изоферменты – ферменты, катализирующие одну и ту же химическую реакцию, но с разной первичной структурой белка. Они отличаются друг от друга кинетическими параметрами, условиями активации, особенностями связи апофермента и кофермента. Возникают изоформы в основном в результате небольших генетических различий в первичной структуре фермента.
Они часто являются органоспецифичными. То есть опреденная изоформа фермента встречается в одном органе, другая – во втором органе.
Например – ЛДГ – лактактдегидрогеназа, катализирующая обратимую реакцию окисления лактата до пирувата (ПВК).
ЛДГ состоит из 4х субъединиц двух типов – М (muscle) и H (heart). Определённая комбинация этих субъединиц как раз и приводит к появлению изоформ.
ЛДГ1 и ЛДГ2 наиболее активны в сердечной мышце и почках. Значит, они состоят преимущественно из субъединиц H.
ЛДГ4 и ЛДГ5 – в скелетных мышцах и печени. Значит, больше субъединиц М.
Такая же ситуация и с креатинкиназой (КК), катализирующей реакцию образования креатинфосфата.
Она уже состоит из 2-ух субъединиц – М (muscle) и В (brain). Итого мы получаем три возможные изоформы: ВВ, ВМ, ММ. Нетрудно догадаться, что изофермент ВВ находится преимущественно в головном мозге, ММ – в скелетных мышцах, а ВМ – в сердечной мышце.
Они, как и изоформы ЛДГ, имеют разную электрофоретическую подвижность.
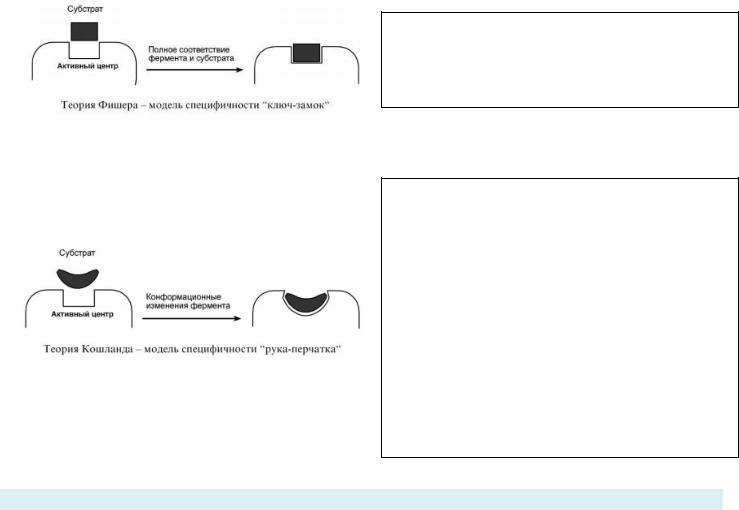
СПЕЦИФИЧНОСТЬ, ВИДЫ СПЕЦИФИЧНОСТИ. МЕХАНИЗМ СПЕЦИФИЧНОСТИ – ТЕОРИЯ ФИШЕРЫ И ТЕОРИЯ КОШЛАНДА:
Специфичность – наиболее важное свойство ферментов, определяющее их биологическую значимость.
Различают субстратную и каталитическую специфичность:
1. Субстратная:
oАбсолютная - фермент производит катализ только одного вещества. Только уреаза может быть ферментом в реакции расщепления мочевины.
oГрупповая - катализ субстратов с общими особенностями. Наблюдаем градацию: субтилизин может разрушить связь между любым аминокислотами, пепсин же катализирует разрыв пептидной связи между аминогруппами только ароматических кислот, то есть он может помочь
разорвать связь между фениаланином, тирозином, триптофаном, а вот тромбин – лишь между аргинином и глицином. Так же алкогольдегидрогеназа может окислить любые OH-группы.
oОтносительная групповая специфичность - превращение субстратов с некоторыми общими признаками. Например, цитохром Р450 окисляет только гидрофобные вещества, которых насчитывается около 7000
2.Каталитическая:
Один и тот же субстрат под действием разных ферментов может преобразоваться в разные продукты.
ГИПОТЕЗА ФИШЕРА: МОДЕЛЬ «ЖЁСТКОЙ МАТРИЦЫ» / «КЛЮЧ-ЗАМОК».
Название говорит само за себя – активный центр фермента идеально соответствует конфигурации субстрата, поэтому, при их соединении, активный центр не меняет свою форму.
ГИПОТЕЗА КОШЛАНДА: МОДЕЛЬ ИНДУЦИРОВАННОГО СООТВЕСТВИЯ / «РУКА-ПЕРЧАТКА».
Активный центр может менять свою форму/конфигурацию, он гибкий. Присоединение субстрата к якорному участку фермента вызывает изменение конфигурации каталитического центра таким образом, чтобы его форма соответствовала форме субстрата. При этом в субстрате уже деформируются некоторые валентные связи и он, таким образом, подготавливается к дальнейшему каталитическому видоизменению, а в молекуле фермента происходят конформационные перестройки. То есть активный центр – это перчатка, а субстрат – твоя рука. Перчатка облегает ладонь, принимая нужную форму.
ОСНОВНЫЕ СВОЙСТВА ФЕРМЕНТОВ. ГРАФИКИ ЗАВИСИМОСТИ СКОРОСТИ ФЕРМЕНТАТИВНОЙ РЕАКЦИИ:
o Действуют в мягких условиях: термолабильность, зависимость от рН o Высокая эффективность (↑ V р-ции 106 - 108 раз)
o Высокая специфичность
o Скорость ферментативной реакции регулируется (наличие активаторов и ингибиторов) o Способность образовыватьм мультиферментные комплексы
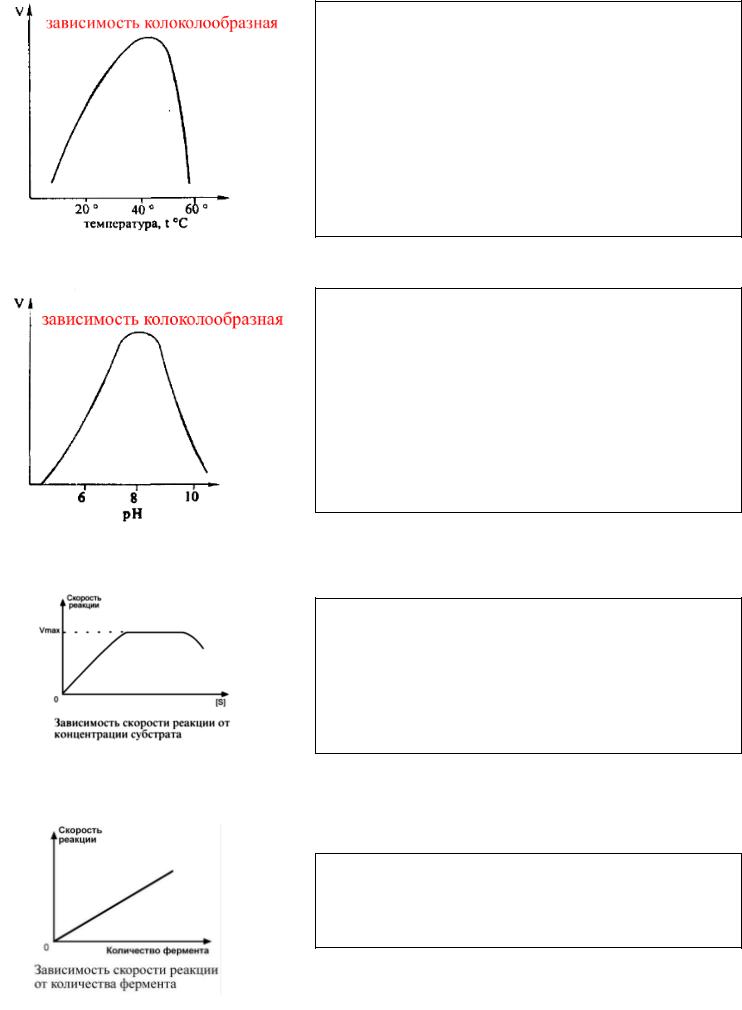
1) Зависимость от температуры:
Наибольшую активность тот или иной фермент проявляет при оптимальной температуре. Для человека это значение находится в пределах +37,0 - +39,0 °С. При понижении температуры, замедляется броуновское движение, уменьшается скорость диффузии и, следовательно, замедляется процесс образования комплекса между ферментом и компонентами реакции (субстратами). В случае повышения температуры выше +40 - +50 °С молекула фермента, которая является белком, подвергается процессу денатурации (именно поэтому там опасна для нас высокая температура). При этом скорость химической реакции заметно падает.
2) Зависимость от РН:
Каждому ферменту комфортно в разных средах, кому-то в кислой, кому-то в щелочной, кому-то в нейтральной. Поэтом график для разных ферментов всегда будет отличаться.
Для многих энзимов оптимум рН 6 - 8, так как сдвиг рН вызывает изменение степени ионизации функциональных групп аминокислот в активном центре СОО- и NH3+.
Оптимальные значения рН для пепсина 1,5-2,5; трипсина 8,0- 8,5; амилазы слюны 7,2; аргиназы 9,7; кислой фосфатазы 4,5- 5,0; сукцинатдегидрогеназы 9,0.
3) Зависимость от концентрации субстрата:
Следим за графиком: сначала концентрация субстрата растёт. Затем наступает эффект насыщения, когда молекул фермента уже просто не осталось. Скорость достигает своего максимума, когда все субстраты связаны с ферментами. А после между молекулами субстрата начинается конкуренция и скорость реакции резко снижается, ну не к чему больше им присоединиться.
4) Зависимость от концентрации фермента:
Концентрация фермента растёт и растёт, прямо пропорционально скорости реакции, так как большее количество молекул фермента производит большее число молекул продукта.

ОСНОВЫ КИНЕМАТИКИ ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ. УРАВНЕНИЕ МИХАЭЛИСА-МЕНТЕН:
Кинетика ферментативных реакций — это раздел энзимологии, изучающий зависимость скорости химических реакций, катализируемых ферментами от химической природы реагирующих веществ и факторов окружающей среды.
Скорость ферментативной реакции является мерой каталитической активности фермента и обозначается как активность фермента.
ТО ЕСТЬ – чем более активный фермент, тем быстрее протекает реакция, то есть увеличивается её скорость.
Эта самая скорость определяется уменьшением количества молекул субстрата или увеличением количества молекул продукта за единицу времени.
ТО ЕСТЬ – чем активнее протекает реакция под действием фермента, тем быстрее субстрат превращается в продукт.
ТО ЕСТЬ – концентрация субстрата уменьшается (он же превращается в продукт), а концентрация продукта растёт (он же образуется, значит, накапливается в клетке).
На практике пользуются условными величинами, характеризующими активность фермента: международная единица активности (ME) соответствует такому количеству фермента, которое катализирует превращение 1 мкмоль субстрата за 1 минуту при оптимальных условиях (температура 37°С, оптимальное значение pH раствора).
Для оценки количества молекул фермента среди других белков данной ткани определяют удельную активность (Уд.Ак.) фермента, численно равную количеству превращенного субстрата (в мкмолях) за единицу времени одним миллиграммом (мг) белка (фермента, выделенного из ткани). По удельной активности судят о степени очистки фермента: чем меньше посторонних белков, тем выше удельная активность.
Если концентрацию фермента оставить постоянной, изменяя только количество субстрата, то график скорости ферментативной реакции описывается гиперболой
О максимальной скорости реакции: это такое состояние, когда все ферменты были связаны со своими субстратами, максимально возможное количество образования продукта. Дальнейшее повышение концентрации субстрата, вроде, должно было повысить скорость образования продукта, но скорость падает из-за конкуренции молекул субстрата.

КОНСТАНТА МИХАЭЛИСА: кинетическая характеристика эффективности фермента. Она численно равна концентрации субстрата, при которой достигается половина максимальной скорости, она характеризует сродство данного фермента к данному субстрату, является величиной постоянной и определяется экспериментально.
Чем меньше Км – тем больше сродство фермента к субстрату, тем выше начальная скорость.
Чем больше Км – тем меньше сродство к субстрату, тем мменьше начальная скорость реакции.
УРАВНЕНИЕ МИХАЭЛИСА-МЕНТЕН:
Уравнение Михаэлиса-Ментен показывает взаимосвязь максимально возможной скорости, реальной скорости реакции, константы Михаэлиса и концентрации субстрата.
1) Концентрация субстрата равна величине констансты Михаэлиса ([S] = Km)
Помнишь, что такое Км? Это концентрация субстрата, при которой скорость реакции равна половине максимальной. Посмотрев на график, получаем, что скорость реакции V будет равна половине максимальной Vmax.(V = ½ Vmax).
2)Концентрация субстрата значительно больше Km ([S] >> Km)
Вэтом случае величиной Km можно пренебречь, при решении получим, что скорость реакции максимальна (плато на графике).
3)Концентрация субстрата значительно меньше Km ([S] << Km).
Вэтом случае, знаменатель уравнения мало изменяется при изменении [S], а величина скорости реакции V прямо пропорциональна [S] (график линеен).