

Ребята, это лекция за 20.03.2020
Задание:
Внимательно прочитать;
Составить конспект;
Если что то совсем не понятно, нужно записать вопрос и задать его любым возможным способом дистанционного обучения;
В конце лекции есть список литературы. Постарайтесь наладить удаленный доступ к электронно-библиотечной системе «Лань». Хорошо бы еще к «Юрайт», но если его еще нет, то удаленно это сделать не получится, а с «Лань» можно попробовать (на ИСУ на главной странице есть инструкция для получения удаленного доступа к «Лань»)
Буду стараться передавать материалы в соответствии с расписанием.
2. 1. ПРЕДМЕТ ЭКОЛОГИИ
Определить предмет экологии позволяет концепция организации живой оболочки 3емли - биосферы. Биосфера состоит из биологических систем - биосистем разного уровня сложности и организации.
Биосистемы - это природные системы, в которых живые компоненты, называемые биотическими, упорядоченно взаимодействуют с неживой физической средой, т. е. с абиотическими компонентами, составляя с ними единое целое. Биосистемы связаны между собой иерархической структурой, образуя как бы «служебную лестницу жизни».
Согласно современным представлениям на «лестнице жизни» выделяют шесть ступенек, которые гипотетически могут быть продолжены и вверх, и вниз (рис. 2.1).

Рисунок 1. Иерархия уровней организации живой материи: 1 - генетические системы; 2- клеточные системы; 3 - системы органов; 4 - системы организмов; 5 - популяционные системы; 6 - экологические системы (экосистемы)
По мере продвижения биосистем от низших ступенек к высшим уровень организации входящих в них биотических компонентов усложняется: гены, клетки, органы, организмы, популяции, сообщества. Абиотические компоненты на всех уровнях представлены веществами и энергией, которые формируют все факторы неживой физической среды.
Биотические и абиотические компоненты вместе образуют соответствующие биосистемы, расположенные снизу вверх в следующем порядке: 1) генетические системы, 2) клеточные системы, 3) системы органов, 4) системы организмов, 5) популяционные системы, 6) экологические системы, или экосистемы.
Деление иерархии на ступени условно, так как каждый уровень взаимосвязан с соседними уровнями в функциональном смысле. Например, гены не могут функционировать в природе вне клетки, клетки многоклеточных - вне органов, органы - вне организма и т. д. Иными словами, системы низших ступеней (подсистемы) интегрированы в системы высших ступеней (надсистемы). Экосистема не жизнеспособна без взаимосвязи с популяционными системами и биосферой в целом. В то же время ни одна из биосистем не может жить, если в ней не происходит обмен веществ и не поступает энергия извне, т. е. отсутствуют абиотические компоненты. Все экосистемы вместе образуют биосферу 3емли.
Биосистемы разных уровней являются предметом изучения различных дисциплин: генетические системы изучает генетиntа (греч. genetikos - наследственный), клеточные - цитология (греч. kytos - клетка), органы - физиология (греч. physis - природа, природные функции). Более высокоорганизованные системы уже не укладываются в рамки одной дисциплины. Организмы, например, рассматривают разнообразные разделы ботаники и зоологии: альгология (лат. alga - водоросль), микробиология (греч. mikros - малый), микология (греч. mykes - гриб), ихтиогия (греч. ichthys - рыба), орнитология (греч. ornis - птица), антропология (греч. antropos - человек) и т. п. Системы, которые расположены выше уровня организмов: популяционные системы, экосистемы, биосферу - изучает экология.
Следовательно, предметом экологии являются биосистемы надорганизменного уровня - популяционные, экологические и биосфера.
2.2. Популяционные системы
Биосистемы, в которых биотические компоненты представлены популяциями, на-
зываются популяционными и характеризуют в основном биологические аспекты экологии.
Популяция - это совокупность разновозрастных особей одного вида, обменивающихся генетической информацией, объединенных общими условиями существования, необходимыми для поддержания численности в течение длительного времени: общностью ареала, происхождением, свободным скрещиванием и др.
В отличие от отдельных организмов популяция характеризуется рядом признаков, носителями которых является группа, но не отдельные особи: плотность, рождаемость, смертность, возрастная структура, распределение в пространстве (дисперсия), кривая роста и др. Кроме «групповых свойств» популяции обладают и «биологическими свойствами», присущими как популяции, так и входящим в нее организмам.
Отдельная особь рождается, стареет, умирает, но применительно к одной особи нельзя говорить о рождаемости, смертности, возрастной структуре, т. е. о характеристиках, имеющих смысл только для группы особей или популяции.
Далее дано описание основных характеристик популяции.
Плотность - это величина численности или биомассы популяции, отнесенная к единице площади или объема пространства. Например, 400 деревьев на 1 км", или 1 млн микроскопических водорослей в 1 м3 воды, или 150 кг рыбы на 1 км2 поверхности водоема.
Важно различать среднюю плотность, т. е. численность или биомассу на единицу всего пространства, и экологическую плотность, т. е. численность или биомассу на единицу фактически обитаемого пространства на территории или акватории, доступной для обитания популяции. Различие между средней и экологической плотностью можно проиллюстрировать рис. 2.2. На рисунке показано, что средняя плотность мелких рыб с падением уровня воды в течение сухого сезона снижается, но экологическая

Рис. 2.2. Экологическая и средняя плотность рыб, служащих добычей аисту в период размножения (по М. Р. Kahe, 1964)
плотность, при этом, возрастает, т. е. по мере сокращения зеркала воды увеличивается число рыб, приходящихся на единицу водной поверхности. Аисты откладывают яйца в такое время, в которое вылупление птенцов приходится на пик экологической плотности рыб. Это облегчает родителям ловлю рыб, которые составляют основную пищу птенцов. Аисты ориентируются на экологическую, а не на среднюю плотность пищевых объектов.
Люди тоже в расчет принимают главным образом пищевую продукцию, сконцентрированную на 15 или 20% поверхности Земли, где возможен высокий урожай, а не на рассеянные и труднодоступные для добывания пищевые ресурсы.
Для измерения плотности популяций существует много разных методов. Но ничто не может заменить непосредственные наблюдения и опыт, приобретенный при полевых исследованиях в природе.
Рождаемость - это способность популяции к увеличению численности. Рождаемость характеризует появление на свет новых особей любых организмов, независимо от того, вылупляются ли они из яиц, прорастают из семян или появляются в результате деления.
Максимальная рождаемость (абсолютная) - это теоретический максимум скорости образования новых особей в идеальных условиях при отсутствии лимитирующих факторов.
Реализованная, или экологическая, рождаемость - это увеличение численности популяции при фактических условиях среды. Эта величина непостоянна и меняется в зависимости от размеров и возрастного состава популяции, а также от физических условий среды обитания.
Рождаемость выражают либо как скорость, определяемую путем деления общего числа появившихся особей на время, - общая рождаемость; либо как число появившихся особей в единицу времени на одну особь в популяции - удельная рождаемость.
Например, популяция из 50 инфузорий в 1 л воды увеличивается путем деления. Через час численность инфузорий возросла до 150 особей. Общая рождаемость равна 100 особей/час, а удельная рождаемость составляет 2 особи/час на 1 исходную особь.
Предположим, что в городе с населением 10 ООО жителей появилось 500 новорожденных. Общая рождаемость составит 500 в год, а удельная - 0,05 или 5%. В демографии принято рассчитывать рождаемость на одну женщину репродуктивного возраста, а не на все население.
Различие между максимальной и экологической рождаемостью можно проиллюстрировать данными, полученными при изучении популяции певчей птицы (Sialis sialis) в городском парке. Птицы отложили 510 яиц (около 15 на одну самку), что соответствует максимальной рождаемости. Оперилось всего только 265 птенцов, что соответствует экологической рождаемости, которая составляет 52% от максимальной, т. е. около 8 птенцов на одну самку.
В опытах на мучном хрущике экологическая рождаемость составила лишь 6% (из 12 ООО яиц вылупилось только 773 личинки).
В общем для видов, которые не охраняют яйца и не заботятся о потомстве, характерна высокая максимальная и низкая экологическая рождаемость, и отношение максимальной к экологической рождаемости очень высокое.
Смертность отражает гибель особей в популяции. Подобно рождаемости, смертность можно выразить числом особей, погибших в единицу времени, или удельной смертностью в расчете на численность популяции. Различают также минимальную и экологическую смертность. Минимальная смертность постоянна для популяции. Она представляет собой гибель особей в идеальных· условиях, так как даже в самых лучших условиях организмы будут умирать от старости.
Возраст, в котором особь умирает от старости, определяется физиологической продолжительностью жизни, которая часто намного превышает экологическую продолжительность жизни. Экологическая, или реализованная, смертность - это гибель особей в реальных условиях среды; она не постоянна и меняется в зависимости от изменения окружающей среды и состояния самой популяции.
Практически большее значение имеет не смертность, а выживаемость организмов в данных условиях.
Как рождаемость, так и смертность, особенно у высших организмов, варьирует с возрастом. Поэтому важно определение удельной смертности для разных возрастных групп или стадий развития.
Распределение выживших особей по возрастным группам изображают в виде кривых выживания, при построении которых на оси абсцисс отложен возраст погибших особей в процентах от физиологической продолжительности жизни, а на оси ординат - число выживших на тысячу особей в логарифмической шкале (рис. 2.3).

Рис. 2.3. Различные типы кривых выживания (по Б. А. :Крискунову, 1995)
Кривые выживания подразделяют на три основных типа. Сильно выпуклая (1) кривая характерна для многих видов крупных животных и человека. Сильно вогнутая кривая (ПI) характеризует организмы с высокой смертностью на ранних стадиях. Например, для двустворчатых моллюсков, рыб, дубов (высокая гибель прорастающих желудей) и др.
Промежуточный тип кривой выживания (П) имеют виды, у которых удельная выживаемость для каждой возрастной группы более или менее одинакова, и в полулогарифмическом масштабе. кривая приближается к диагонали.
Форма кривой выживания связана со степенью заботы о потомстве. Так, кривые выживания пчел (заботятся о потомстве) значительно менее вогнуты, чем у кузнечиков (не заботятся о потомстве). :Кузнечики компенсируют гибель особей ранних возрастных групп большим числом откладываемых яиц, и отношение максимальной и экологической рождаемости у них очень велико.
Форма
кривой выживания часто варьирует в
зависимости от плотности популяции.
Например, кривая выживания чернохвостого
оленя, обитающего в Калифорнии, при
плотности 40 оленей на 1 км2 значительно
более вогнута, чем кривая

Гидра -

выживаемости тех же оленей при их плотности 1 7 оленей на 1 км2• Вогнутость кривой выживания при высокой плотности оленей может быть связана с интенсивной охотой и внутривидовой конкуренцией.
Кривая выживания человека приближается к остроугольному типу (1). И хотя экологическая продолжительность жизни человека, благодаря медицине и питанию, увеличилась, физиологическая продолжительность жизни при этом практически не изменилась по сравнению с предыдущими столетиями - редко кто доживает до 100 лет. Возможно, это связано с тем, что высокая плотность населения в популяциях человека опасна для качества, жизни отдельного индивидуума.
Пространственная структура популяций, т. е. характер распределения организмов в пространстве, может быть: случайным, равномерным и групповым (рис. 2.4).

Рис. 2.4. Три основных типа распределения особей в пространстве:
А- случайное; Б - равномерное; В- групповое (по Ю. Одуму, 1986)
Случайное распределение наблюдается тогда, когда среда обитания очень однородна, и организмы не стремятся объединяться. Равномерное встречается там, где между особями сильная конкуренция или существует антагонизм другого типа. Но чаще в популяции наблюдается тенденция к образованию групп особей (пары, семьи, стаи, стада). Распределение самих групп может быть случайным или равномерным. Тип распределения особей важно учитывать при выборе метода взятия проб и учета численности организмов на экспериментальных площадках: число выборок для расчетов численности при случайном и групповом распределении должно быть больше, чем при равномерном. При малой выборке из популяции с групповым распределением будут получены либо слишком низкие, либо, слишком высокие результаты при пересчете на всю популяцию.
Таким образом, для большинства популяций характерно образование групп разных размеров в разное время. Агрегирование особей в группы происходит по разным причинам: вследствие различий в местообитаниях; под влиянием суточных и сезонных изменений погоды; в связи с процессами размножения и у высших животных стремлением к социальному объединению. Жизнеспособность группы по сравнению с одиночками, несмотря на возникающую иногда конкуренцию, повышается. Группа обладает большими возможностями для защиты, добычи ресурсов или изменения микроклимата и микроместообитания. Степень агрегации, при которой наблюдается оптимальный рост и выживание популяции, варьирует у разных видов в разных условиях.
Отсутствие агрегации ( «недонаселенностъ» ), так же как и чрезмерная агрегация («перенаселенностъ»), могут тормозить оптимальное развитие популяции. Наиболее благополучная популяция характеризуется промежуточными размерами агрегации (плотности). Эта закономерность получила название принципа Олли (рис. 2.5). Принцип Олли вполне приложим к человеку. «Городская агрегация» благоприятна для человека до определенного предела.

Рис. 2.5. Иллюстрация принципа Олли (по Allee et al., 1949)
Если построить график зависимости выгоды (ось у) от величины города (осъ х) , то он будет иметь такой же вид, как кривая -- на рис. 2.5. Население городов следует уменьшать, как только стоимость жизни в городе начнет превышать выгоду с точки зрения экологии. Ошибочно субсидировать город, который стал слишком большим для своего обеспечения.
Особый тип агрегации называется образованием безопасных поселений. В этом случае социально организованные группы животных поселяются в благоприятно расположенном участке, откуда расходятся для удовлетворения пищевых и других потребностей и куда возвращаются. Человек также использует эту стратегию. В результате агрегации повышается выживаемость, особенно у животных.
Например, Олли обнаружил, что рыбы в группе лучше переносят дозы яда, введенные в воду, чем изолированные особи. Слизь и другие выделения группы рыб способствуют противодействию ядам. Пчелы в улье также выделяют и сохраняют достаточно тепла для выживания всех особей при внешней температуре, при которой гибнут изолированные насекомые.
Колониальные птицы часто не могут размножаться, если колония становится маленькой. Олли отметил, что даже у примитивных организмов встречается кооперация, и это начало социальной организации, в разной степени развитой в животном мире и достигающей кульминации у человека.
Очень высокий уровень специализации наблюдается у общественных насекомых (термиты, муравьи, пчелы). Разделение труда у них обеспечивается существованием трех каст: производителей, рабочих и солдат. Каждая из каст морфологически специализирована для соответствующих функций: размножения, заготовки пищи и защиты.
Настоящая социальная агрегация характеризуется определенной организацией, социальной иерархией и специализацией особей (рис. 2.6).

Рис. 2.6. Порядок стада павианов в пути (А) и при нападении леопарда (В):
В - вожаки; Н - самцы низшего порядка; С - самки; М - молодняк (по Черновой, Быловой, 1988)
Возрастная структура популяции является одной из самых важных ее характеристик, которая влияет на рождаемость и смертность. Соотношение разных возрастных групп определяет способность популяции к размножению. В быстрорастущих развивающихся популяциях значительная доля приходится на молодые особи; в популяциях, находящихся в стабильном состоянии, возрастное распределение относительно равномерно, а в отмирающих популяциях молодые особи составляют меньшую долю от общей численности (рис. 2. 7).

Рис. 2.7. Возрастная структура популяций
Для каждой популяции характерно «нормальное» или стабильное распределение возрастных групп. Отклонения от этого состояния могут быть вызваны взрывом рождаемости или смертности, после чего происходит спонтанный возврат к стабильной ситуации. В человеческом обществе также по мере его развития происходит переход от быстрого роста численности к зрелому стабильному состоянию. При этом процент младших возрастных групп уменьшается и увеличивается процент населения старшего возраста, что оказывает большое воздействие на жизненный уклад общества и его экономику.
В популяциях обычно выделяют три возрастные группы: пререпродуктивную, репродуктивную и пострепродуктивную. Длительность этих возрастов сильно варьирует у разных организмов. У современного человека продолжительность этих «возрастов» приблизительно одинакова, т. е. около трети жизни. У первобытных людей пострепродуктивный возраст был намного короче. У некоторых бабочек, например, пререпродуктивный период очень длительный, репродуктивный - короткий, а пострепродуктиввый очень короткий или может вообще отсутствовать. Возрастная структура популяций зависит от особенностей размножения и продолжительности жизни особей.
Анализ возрастной структуры позволяет прогнозировать численность популяций на ряд ближайших лет. Подобные прогнозы применяются для оценки уловов промысловых рыб, в охотничьем хозяйстве и других областях сельского хозяйства.
Такие показатели, как рождаемость, смертность, возрастная структура, очень важны, но по каждому из них в отдельности нельзя судить о росте численности популяции как целого, о биотическом потенциале популяции; т. е: «врожденному» свойству организмов к размножению и выживанию, который приводит к увеличению численности.
Популяции изменчивы по своей сути. Все характеристики популяции изменяются по мере того, как вид подстраивается к сезонам, физическим факторам и другим организмам. Поэтому в большей мере, чем величина и структура популяции в некоторый момент времени, должно интересовать то, как часто и с какой скоростью в популяции происходят изменения. Характер увеличения численности популяций может быть различным, что отражается на форме кривых роста численности.
Кривые роста численности популяций, построенные в арифметическом масштабе, делят на два основных типа: описываемый J-образной и S-образной, или сигмоидной, кривой.
В первом случае численность популяции увеличивается с возрастающей скоростью. При втором типе рост популяции вначале увеличивается медленно, затем быстрее, пока сопротивление среды постепенно не замедлит роста.
Если число организмов обозначить N, время - t, ∆N - изменение числа организмов, то ∆N / ∆t - средняя скорость изменения численности за время t. При этом ∆N /(N∆t)- средняя скорость изменения числа организмов в расчете на одну особь. Этот параметр называется удельной скоростью роста и используется при сравнении популяций.
Умножив удельную скорость на 100, получим скорость роста популяции в процентах. Эти величины можно выразить в дифференциальной форме: dN / dt - скорость изменения числа организмов за единицу времени в некоторый момент.
При J-образной кривой, называемой кривой экспоненциального роста, численность быстро возрастает по экспоненте:
dN/dt=rN,
где константа r характеризует скорость процесса.
Параметр r можно рассматривать как коэффициент мгновенного роста. Это фактически разность между удельной мгновенной скоростью рождаемости а и мгновенной удельной скоростью смерти b:
r=a–b.
Скорость роста популяции (r) в нелимитирувмой среде ( если среда никак не ограничивает рост пространством, пищей, конкурирующими организмами и др.) зависит от возрастного состава и удельной скорости роста возрастных групп.
Когда устанавливается стабильное возрастное распределение, удельную скорость роста популяции называют внутренней или максимальной скоростью роста (r max), которую часто называют еще биотическим или репродуктивным потенциалом.
Разницу между максимальным биотическим потенциалом (rmax) и реальной скоростью роста (r) используют как меру сопротивления среды, которая характеризует сумму всех лимитирующих факторов среды, препятствующих реализации биотического потенциала.
Биотический потенциал - это скрытая потенциальная генетическая способность к размножению в популяциях, которая, к счастью, никогда не реализуется благодаря сопротивлению среды. В противном случае в нелимитируемых условиях среды потомство только одной пары мух через несколько лет могло бы весить больше, чем земной шар.
При сигмоидной кривой рост популяции вначале увеличивается медленно (фаза положительного ускорения), затем быстро (приближается к логарифмической фазе), но под влиянием сопротивления среды замедляется (Фаза отрицательного ускорения). Затем будет достигнуто равновеcиe, которое и сохраняется. ·
Такой тип роста описывается логистическим уравнением
dN /dt = rN (K-N)/K,
где К - верхний предел увеличения численности популяции, называемый верхним пределом роста.
При S-образной кривой рост численности называется логистическим (рис. 2.8).
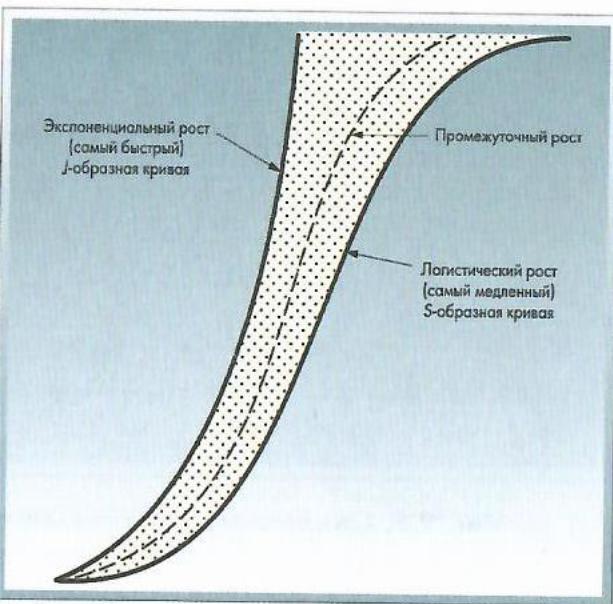
Рис. 2.8. Экспоненцнальвая и логистическая кривая роста численности (по Wiegert, 1974)
Рост большинства популяций занимает промежуточное положение.
J-образную кривую можно рассматривать как неполную сигмоидную кривую, в случае которой лимитирующие факторы среды начинают ограничивать рождаемость до того, как начнут действовать внутренние факторы ограничения численности.
Когда популяция завершает рост и средняя скорость роста за длительное время под воздействием внешних и внутренних факторов становится равной нулю, численность ее резко падает, но затем восстанавливается.. Популяция проявляет тенденцию к колебаниям численности относительно стационарного уровня. Такие колебания, как правило, обусловлены сезонными или годовыми колебаниями условий среды (температура, осадки, доступность пищи или энергии, болезни и др.).
Всем хорошо известны сезонные изменения численности и плотности популяций: тучи комаров; леса, полные птиц; поля, заросшие ромашками или васильками, - все это бывает в свой сезон - в другое время популяции этих организмов могут сходить на нет.
У некоторых популяций колебания численности столь регулярны, что являются «циклическими», Классическим примером 9-10-летних колебаний служит изменение численности зайца-беляка и рыси (рис. 2.9).

Рис. 2.9. Циклические колебания численности зайца-беляка и рыси (по Крискунову и др., 1995
У рыси пики численности повторяются каждые 9-10 лет (наблюдения велись с 1800 г.), которые сменяются резкими спадами, когда в течение нескольких лет рысь встречается крайне редко. У зайца-беляка пики численности на год или больше предшествуют пикам численности рыси. Поскольку рысь питается зайцами, ясно, что цикл хищника связан с циклом жертвы. Но циклы численности зайца встречаются и в тех местах, где рысь не водится. В этом случае численность популяции может регулироваться у многих млекопитающих перенаселением, вызывающим изменение
физиологического состояния, приводящего к болезням и другим стрессам. Зайцы-беляки, например, в период пика численности часто внезапно погибают от «шоковой болезни».
Такие механизмы относятся к внутренним регуляторам численности и срабатывают автоматически, когда плотность превысит некоторое пороговое значение.
Для оценки причин изменения численности растений и животных и влияния человека на окружающую природную среду необходимы знания не только о свойствах отдельных организмов, но о видовых популяциях - основных функциональных единицах природы.
Популяции - элементы экосистем и участники процессов трансформации веществ и переноса энергии. Популяции постоянно изменяются и находятся в динамике. Динамика популяции - результат совместного действия множества факторов окружающей природной среды и внутренних механизмов регуляции всех основных ее характеристик.