

9654
.pdf71
это бывает, когда почва хорошо прогрета и корни активно всасывают воду,
а транспирация отсутствует (например, утром или при тумане, когда
влажность |
воздуха |
100%). |
Избыточная |
влага |
удаляется путем |
г у т т а ц и и |
(рис. |
24, 25) – |
выделения |
воды |
через специальные |
выделительные клетки, расположенные по краю или на острие листа. К
гуттации способны не только гигрофиты, но и многие мезофиты. Так, в
украинских степях гуттация обнаружена более чем у половины всех видов.
Многие луговые травы гуттируют столь сильно, что увлажняют поверхность почвы. По словам А. Фрей-Вислинга (1949), при более точном наблюдении можно установить, что большая часть росы, увлажняющей по утрам наши луга, не падает с неба, а выделяется самими луговыми травами.
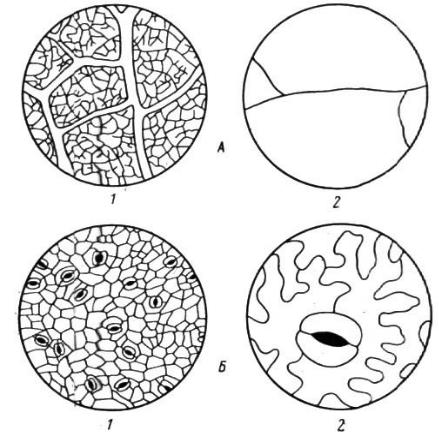
Рис. 26. Рыхлые ткани с системой межклетников на поперечных срезах прибрежного растения Alisma plantago-aquatica (по Василевской В. К., из Шенникова А. П., 1950): А – лист; Б – стебель; В – корень
Избыточное увлажнение почвы может сказываться на растениях и отрицательно, поскольку затрудняет аэрацию, а, следовательно, дыхание и всасывающую деятельность корней. Борьба растений с избыточной влагой
– это, прежде всего, борьба за доступ воздуха. У многих травянистых видов избыточно увлажненных местообитаний (прибрежных, болотных) хорошо развита система воздушных полостей в листьях и стеблях, а также специальная воздухоносная ткань – аэренхима (рис. 26). Благодаря этому создается непрерывный путь от надземных частей к корням, по которому
72
подземные части растений снабжаются воздухом. Так, экспериментально показано передвижение кислорода вниз по стеблям прибрежных трав
(манника Glycerin aquatica, камыша Scirpus sylvaticus и др.), у которых стебель представляет собой полую трубку со стойкими стенками.
Влияние избытка влаги в почве испытывают и леса. На севере и северо-
западе европейской части России заболоченные леса и насаждения с посто-
янным или временным (весной и осенью) избытком влаги занимают около
60% всех лесных площадей. В таких местообитаниях на древесные породы угнетающе действует недостаток или полное отсутствие в воде свободного кислорода, а также появление недоокисленных соединений, накопление токсических веществ – СО2, H2S и др., что приводит к снижению поглощения корнями влаги и минеральных веществ из почвенного раствора, затруднению аэробного дыхания корней, сокращению фотосинтеза и другим нарушениям.
Ксерофиты – это растения сухих местообитаний, способные пе-
реносить значительный недостаток влаги – почвенную и атмосферную засуху. Ксерофиты распространены в областях с жарким и сухим климатом. Они обильны и разнообразны. К этой группе принадлежат виды пустынь, сухих степей, саванн, колючих редколесий, сухих субтропиков и т. д. В более гумидных районах ксерофиты участвуют в растительном покрове лишь в наиболее прогреваемых и наименее увлажненных местообитаниях (например, на склонах южной экспозиции, на скалах и т. д.).
Неблагоприятный водный режим растений в сухих местообитаниях обусловлен: во-первых, ограниченным поступлением воды при недостатке в почве и, во-вторых, увеличением расхода влаги на транспирацию при большой сухости воздуха и высоких температурах. Следовательно, для преодоления недостатка влаги возможны разные пути: увеличение ее поглощения и сокращение расхода, кроме того, способность переносить
73
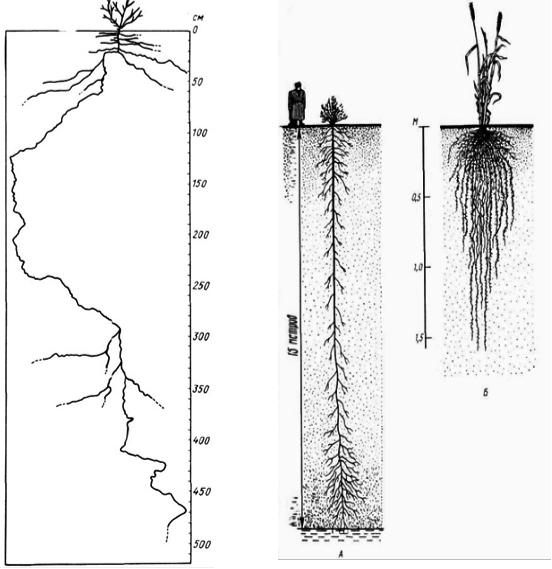
большие потери воды. Все это используется ксерофитами при адаптации к сухости, но у разных растений в неодинаковой степени, в связи с чем некоторые авторы различают два основных способа преодоления ксерофитами засухи: возможность противостоять иссушению тканей или активно регулировать водный баланс и способность выносить сильное иссушение. Большое значение для ксерофитов имеют разнообразные структурные приспособления к условиям недостатка влаги. Корневые системы обычно сильно развиты (рис. 27), что помогает растениям увеличить поглощение почвенной влаги. По общей массе корневые системы ксерофитов нередко превышают надземные части, иногда весьма значительно. Так, у многих травянистых и кустарниковых видов среднеазиатских пустынь подземная масса больше надземной в 9–10 раз, а у ксерофитов памирских высокогорных холодных пустынь – в 300–400 раз.
Корневые системы ксерофитов часто бывают экстенсивного типа (рис. 28) ,
т. е. растения имеют длинные корни, распространяющиеся в большом объеме почвы, но сравнительно мало разветвленные. Проникновение таких корней на большую глубину позволяет ксерофитам (например, многим пустынным кустарникам) использовать влагу глубоких почвенных горизонтов, а в отдельных случаях – и грунтовых вод. Встречаются и широко раскинутые поверхностные корневые системы, приспособленные к поглощению скудных атмосферных осадков, смачивающих лишь верхние горизонты почвы.
У других видов (например, степных злаков) корневые системы интенсивного типа (рис. 28): они охватывают относительно небольшой объем почвы, но благодаря очень густому ветвлению максимально используют почвенную влагу. Корни ряда ксерофильных видов имеют специальные приспособления для запасания влаги («корневые шишки»). Они присутствуют у степной таволги Filipendula hexapetala, получившей из-за этого название «земляные орешки»; утолщенные и деформированные корни пустынных кустарников из рода Calligonutn. Надземные органы ксерофитов
74
также отличаются своеобразными (так называемыми ксероморфными)
чертами, которые носят отпечаток трудных условий водоснабжения. У них сильно развита водопроводящая система, что хорошо заметно по густоте сети жилок в листьях (рис.29), подводящих воду к тканям. Эта черта облегчает ксерофитам пополнение запасов влаги, расходуемой на транспирацию.
Рис. 27. Корневая система |
Рис. 28. Различные типы корневых |
пустынного кустарника Pitiranthos |
систем. |
tortuosus (по Вальтеру Г., 1968) |
А – экстенсивный (верблюжья колючка |
|
Alhagi camelorum); Б – интенсивный |
|
(пшеница) |
75
Ниже показана густота жилкования в листьях растений различных
местообитаний (по данным разных авторов):
Polygonatum multiflorum………………………………………… |
250 |
Тундровые кустарнички: |
|
Betula nana………………………………………………………. |
690 |
Dryas punctata…………………………………………………… |
220 |
Salix polaris……………………………………………………… |
180 |
Пустынный кустарник – Rosa petsica……………………………… |
3130 |
Рис. 29. Различие жилкования (А), размеров по числу устьиц (Б) у ксерофитов и мезофитов (по Келлеру Э. Ф., из Шенникова А. П., 1950):
пустынный ксерофит – Psoralea drupacea, 2 – лесной мезофит – Paris quadrifolia
Разнообразные структурные приспособления защитного характера,
направленные на уменьшение расхода воды, в основном сводятся к следующему:
1. Общее сокращение транспирирующей поверхности. Многие ксерофиты имеют мелкие, узкие, сильно редуцированные листовые пластинки. Это особенно наглядно при сравнении облика родственных
76
видов из сухих и влажных местообитаний. Достаточно вспомнить,
например, мелкие рассеченные листья степной таволги Filiperdula hexapetala и крупные листовые пластинки лабазника Filipendula ulmaria,
обитающего на сырых лугах и опушках; узкие проволоковидные листья степного типчака Festuca sulcata и листья луговой овсяницы Festuca pratensis и т. д. В особо засушливых, пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцированы до едва заметных чешуек. У таких афилльных (безлистных) видов фотосинтез осуществляют зеленые ветви (например, саксаулы Haloxylon aphyllum, Haloxylon persiсит и др., кустарниковые солянки из рода Salsola, кандым
Calligonum, хвойник Ephedra и т. д.). По данным Г. Вальтера, в аридных областях в результате сильного сокращения общей испаряющей массы растительного покрова на единицу количества выпадающих осадков приходится примерно столько же транспирирующей поверхности, сколько в гумидных областях, и в целом расходы воды на единицу листовой площади довольно близки.
2. Уменьшение листовой поверхности в наиболее жаркие и сухие периоды вегетационного сезона (рис. 30). Для многих кустарников среднеазиатских, североафриканских и других пустынь, а также для некоторых видов сухих субтропиков Средиземноморья характерен сезонный диморфизм листьев: ранней весной при еще благоприятном водном режиме образуются относительно крупные листья, которые летом,
при наступлении жары и сухости, сменяются мелкими листьями более ксероморфного строения с меньшей интенсивностыо транспирации. У
деревьев в тропических областях с ярко выраженным сухим периодом распространен летний листопад.
3.Защита листьев от больших потерь влаги на транспирацию.
Она достигается благодаря развитию мощных покровных тканей —
толстостенного, иногда многослойного эпидермиса, часто несущего

77
различные выросты и волоски, которые образуют густое «войлочное» опушение поверхности листа. Сильно опушенные листья имеют степные ксерофиты: коровяк, или «медвежье ухо» Verbascum thapsis, вероника
Veronica incana, котовик Nepeta cataria, многие полыни, шалфеи и др. У
других видов поверхность покрыта водонепроницаемым слоем толстой кутикулы или воскового налета. Развитие защитных покровов на листьях – причина того, что степной травостой имеет тусклые, седоватые оттенки,
резко отличающиеся от яркой зелени лугов.
Рис. 30. Морфологические типы растений засушливых областей (по Вальтеру Г.,
1968):
1 – эфемероиды; 2 – Lycium; 3 – Zygophyllum; 4 – Retama; 5 – Ethedra; 6 – Artemisia; 7 – Reaumuria; 8 – Noaea; 9 – Anabasis; 10 – Haloxylon persicum. Штриховка – ассимилирующие части, отмирающие при недостатке влаги; белый цвет – части, сохраняющиеся при засухе; точки – неассимилирующие части
78
Устьица у ксерофитов обычно защищены от чрезмерной потери влаги, например, расположены в специальных углублениях в ткани листа
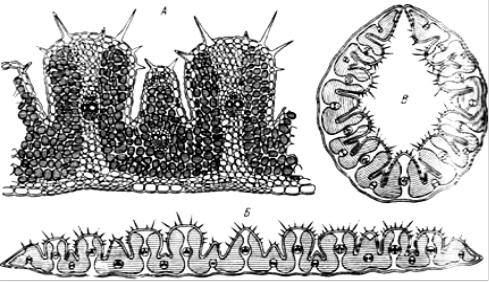
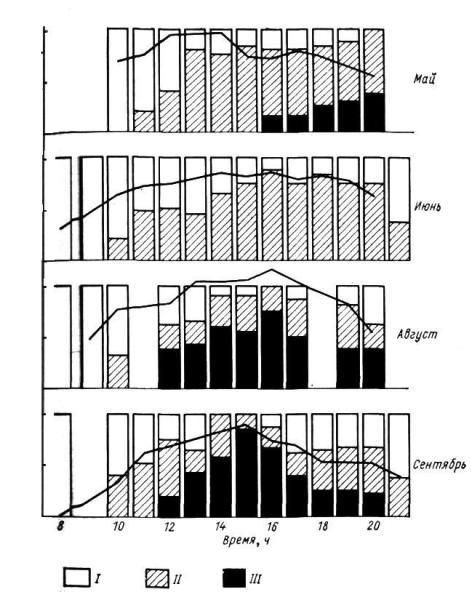
(«погруженные устьица»), иногда снабжены волосками и прочими дополнительными защитными устройствами. У ковылей и других степных злаков существует интересный механизм защиты устьица в самые жаркие
исухие часы дня: клетки эпидермиса теряют тургор, и лист свертывается в трубку (рис. 31, 32). Так, устьица оказываются изолированными от окружающего сухого воздуха внутри замкнутой полости, где благодаря транспирации создается при больших потерях воды повышенная влажность. Во влажную погоду клетки эпидермиса восстанавливают тургор
илистовая пластинка вновь развертывается.
Рис. 31. Поперечный срез листа ковыля-волосатика – Stipa capillaSa (по Кернеру А.,
1896):
А – участок среза при большом увеличении; Б – во влажную погоду (пластинка листа развернута); В – при засухе (лист свернут)
4. Усиленное развитие механической ткани, которое некоторые авторы рассматривают как приспособление, предупреждающее обвисание листовых пластинок при больших потерях воды. Клетки тканей листьев у ксерофитов отличаются мелкими размерами и весьма плотной упаковкой,
т. е. малым развитием межклетников, благодаря чему сильно сокращается внутренняя испаряющая поверхность листа. Поскольку ксерофиты обычно
79
обитают на открытых, хорошо освещенных местообитаниях, многие черты ксероморфной структуры листа – это одновременно и черты световой структуры. Так, у многих видов листья имеют мощную, иногда многорядную палисадную паренхиму, часто расположенную с обеих сторон
(изопалисадные листья) (рис. 33). Формирование некоторых черт ксероморфной структуры (в частности, мелких размеров клеток) – прямое следствие недостатка воды, тормозящего растяжение клеток в растущем листе.
С
30
0
0
0
0
Рис. 32. Дневные ритмы свертывания листовых пластинок ковыля — Stip rubens в казахстанских степях (по Свешниковой В. М. и Бобровской Н. И., 1973): Пластинки: I – полностью раскрыты, II – полусвернуты, III – свернуты. Кривая – дневной ход температуры воздуха
80
Ксерофиты с наиболее ярко выраженными ксероморфными чертами строения листьев, перечисленными выше, имеют своеобразный внешний облик, за что получили название склерофитов. Облик типичного склерофита легко представить на примере чертополоха Carduus crispus,
степных и пустынных полыней, ковылей, саксаулов.
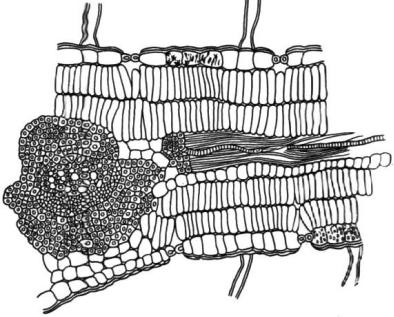
Рис. 33. Поперечный срез листа пустынной древесной породы – песчаной гкалии
Ammodendron conollyi (по Шенникову А. П., 1950)
Разнообразны физиологические адаптации ксерофитов, помогающие им успешно выдерживать недостаток влаги. У ксерофитов обычно повышено осмотическое давление клеточного сока, позволяющее всасывать воду даже при больших водоотнимающих силах почвы, т. е.
использовать не только легкодоступную, но и труднодоступную почвенную влагу. Оно измеряется тысячами килопаскалей, а у некоторых пустынных кустарников зарегистрированы цифры, достигающие 10 000– 30 000 кПа.
С давних пор пристальное внимание привлекала проблема расхода воды ксерофитами на транспирацию. Казалось бы, многочисленные