

Другие файлы / Методы оценки размеров популяции
.docxМетоды оценки размеров популяции
При проведении любых количественных экологических исследований важно с большой степенью точности дать оценку численности организмов, населяющих определенную площадь на суше или объем воды или воздуха. Как правило, это равнозначно оценке величины популяции. Выбор метода зависит от размеров и образа жизни организма и площади исследуемой территории. На небольшом участке можно непосредственно подсчитать число или оценить проективное покрытие или обилие растений и прикрепленных или медленно передвигающихся животных. На больших же открытых пространствах для оценки численности быстро передвигающихся животных необходимы косвенные методы учета. В местообитаниях, в которых наблюдение за организмами затруднено вследствие особенностей их поведения и образа жизни, приходится оценивать численность организмов, используя либо метод изъятия, либо метод мечения и повторного отлова. Существуют объективные и субъективные методы оценки численности популяции.
Объективные методы
Квадраты, прямое наблюдение и фотографирование относятся к методам прямого учета, тогда как метод изъятия и повторного отлова организмов составляют косвенные методы учета.
Квадрат. Если установлено число организмов в пределах некоторого числа квадратов, занимающих известную часть общей площади, то простым умножением можно подсчитать численность организмов на всей территории. Используя этот метод, можно определить следующие три параметра распределения видов.
1) Плотность вида. Плотность вида - это число организмов определенного данного вида на данной площади, например на 10м2. При определении плотности подсчитывают число организмов в брошенных случайным образом квадратах. Метод имеет ряд преимуществ, а именно: он точен, позволяет сравнивать различные участки и виды и установить точное значение обилия. Недостатки метода состоят в том, что он требует значительных затрат времени и при пользовании им необходимо четко определить категорию "особь". Например, считать за одно растение всю дернину или каждый побег?
2) Частота вида. Это вероятность обнаружения определенного вида в пределах любого случайным образом брошенного квадрата в данном районе. Например, если вид встречается в одном из каждых десяти квадратов, то его встречаемость равна 10%. Чтобы получить эту величину, отмечают наличие или отсутствие вида в каждом квадрате, брошенном случайным образом. (Число присутствующих особей не имеет значения.) При использовании этого метода необходимо оговорить размер квадрата, так как это будет влиять на результаты, и, кроме того, условиться о том, какую частоту учитывать: побеговую или корневую? (В случае "побеговой" частоты вид считается присутствующим, если его листья попадают на площадь квадрата, хотя сама особь находится вне его. При "корневой" встречаемости вид считается присутствующим, если его особи укореняются на площади, захваченной квадратом.) Преимущества этого метода в том, что он прост и не требует больших затрат времени. Его применяют в определенных крупномасштабных экосистемах, например в лесных массивах. Недостатки же состоят в том, что на полученное значение частоты влияют размеры квадрата, размеры растения и его пространственное размещение (например, случайное, равномерное или групповое).
3) Проективное покрытие вида показывает, какая часть почвы занята особями данного вида, и дает оценку площади, покрытой этим видом в процентах от общей площади. Его определяют в нескольких случайно выбранных точках путем регистрации покрывающего почву вида, каждый раз субъективно оценивая площадь квадрата, покрытую этим видом, или используя раму со стержнем (рис. 13.13). Этот метод удобен при оценке проективного покрытия растений, особенно травянистых, когда посчитать число особей трудно и не столь важно, как определить проективное покрытие. Однако это достаточно трудоемкий и утомительный метод.
Прямое наблюдение. Прямой подсчет особей применим не только к сидячим или медленно передвигающимся животным, но и ко многим крупным подвижным организмам. К таким, например, как олени, дикие пони и львы, лесные голуби и летучие мыши, в то время когда они покидают место ночлега.
Фотографирование. Прямым подсчетом особей на фотоснимках можно установить размеры популяций крупных млекопитающих и морских птиц, собирающихся на открытых пространствах.
Метод изъятия. Этот метод очень удобен для оценки численности мелких организмов, особенно насекомых, на определенном участке луга или в определенном объеме воды. Взмахами специальной сетки животных отлавливают, записывают число пойманных и не выпускают до конца исследования. Затем еще трижды повторяют отлов, при этом с каждым разом число пойманных животных уменьшается. При построении графика отмечают число пойманных при каждом отлове животных против общего числа пойманных ранее животных. Продолжив линию графика к точке, в которой вновь пойманных животных не окажется (т.е. их число в последнем отлове равно нулю), можно оценить общий размер популяции, как это показано на рис. 13.14.
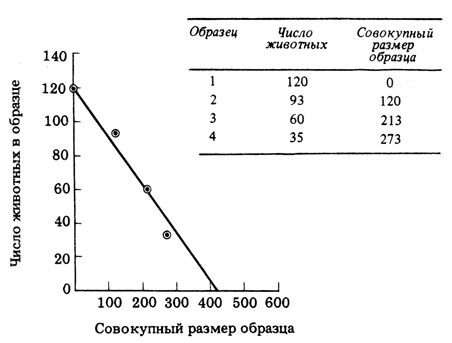
Рис. . График зависимости числа животных в каждом улове от общего числа пойманных ранее животных. Продолжив линию к точке, в которой число вновь пойманных животных равно О, получим оценку численности популяции
Метод мечения и повторного отлова. Этот метод включает отлов животного, его мечение таким образом, чтобы не причинить ему вреда и выпуск на волю там, где его поймали, с тем чтобы оно могло продолжить нормальную жизнедеятельность в популяции. Например, на жаберные крышки пойманной сетями рыбы прикрепляют алюминиевые пластинки или на ноги пойманных сетями птиц надевают кольца. Мелких млекопитающих можно метить краской, надрезать ухо или остригать пальцы, членистоногих также метят краской. В любом из случаев можно применить такую форму кодирования, которая позволяет различить индивидуальные организмы. Пойманных животных подсчитывают, метят репрезентативную выборку из них, затем всех животных выпускают в то же самое место. Через некоторое время животных снова отлавливают и подсчитывают в выборке число животных с меткой. Размер популяции оценивают, используя следующее словесное уравнение:
Общий размер популяции = Число животных в первом улове х Число животных во втором улове
Число животных с меткой во втором улове
Эта оценка размера популяции называется индексом Линкольна. Индекс выводится исходя из ряда приведенных ниже допущений.
1. Внутри популяции организмы размещены случайным образом. (Это не всегда так, поскольку некоторые организмы живут колониями, стадами или стаями.)
2. Между первым и вторым отловом должно пройти достаточное количество времени, чтобы животные успели распределиться случайным образом. Чем менее подвижны особи вида, тем больше времени должно пройти между отловами.
3. Индекс применим только к популяциям, перемещение которых ограничено по географическим причинам.
4. Организмы равномерно размещены в пределах географического ареала популяции.
5. Изменения размеров популяции, вызываемые иммиграцией, переселением, рождением и смертью особей, незначительны.
6. Метки не должны мешать передвижению животных и делать их заметными для хищников.
При изучении растений и мелких животных, например усоногих раков, прямой подсчет численности - очень трудоемкая процедура, поэтому ее в зависимости от необходимой степени точности можно заменить оценкой проективного покрытия или обилия на площади квадратной рамы. В начале работы, для того чтобы увеличить точность оценки, рекомендуется пользоваться рамой, поделенной на мелкие квадратики (рис.). Для определения проективного покрытия или обилия можно освоить различные методики, одни из которых целиком субъективны, другие частично или полностью объективны.
Субъективные методы
Эти методы включают в себя определенный способ оценки частоты, шкалу частоты или оценку обилия через покрытие. Например, в условной шкале, составленной Криспом и Саутвардом, для оценки обилия живущих на скалистых берегах блюдечек (Patella) используются следующие символы, частоты и проценты.
А-обильный > 50%
С-обычный 10-50%
F-часто встречающийся 1-10%
О - редкий < 1 %
R - единичный - за 30 минут поисков обнаружено лишь несколько особей
Соотношение процентов и словесных обозначений частоты условное, поэтому оно может меняться. Например, в отдельных случаях обильным может считаться вид с покрытием > 90%. Значения приведенных выше пяти категорий можно использовать при представлении данных, например при построении кайт-диаграмм (разд. П.2.7.3). Главный недостаток этого метода состоит в том, что он необъективен и при использовании его наблюдается тенденция к уменьшению величины покрытия для мелких невзрачных видов по сравнению с цветущими, бросающимися в глаза и растущими в куртинах видами.
Численность популяции не произвольна даже при постоянной средней продолжительности жизни, а изменяется в пределах определенного диапазона. В соответствии с правилом максимума размера колебаний плотности популяционного населения Ю. Одума,
существуют определенные верхние и нижние пределы для размеров плотности (численности) популяции, которые соблюдаются в природе или которые теоретически могли бы существовать в течение сколь угодно длительного отрезка времени в условиях стабильности среды обитания.
К. Фридерихсом (1927) была сформулирована теория биоценотической регуляции численности популяции:
регулирование численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида.
Размеры популяции возрастают в результате иммиграции из соседних популяций и (или) за счет размножения особей.
Общая численность и плотность населения популяций регулируется правилом максимальной рождаемости (воспроизводства):
в популяции имеется тенденция к образованию теоретически максимально возможного количества новых особей.
Максимальную рождаемость достигают в идеальных условиях, когда отсутствуют лимитирующие экологические факторы, и размножение ограничено лишь физиологическими особенностями вида. Обычно существует экологическая или реализуемая рождаемость, возникающая в обычных или специфических условиях среды.
Возрастная и половая структуры популяций
Средняя продолжительность жизни организмов и соотношение численности (или биомассы) особей различного возраста характеризуются возрастной, а соотношение особей разного пола — половой структурами популяции. Формирование возрастной структуры происходит в результате совместного действия процессов размножения и смертности.
Со временем даже в пределах одной и той же популяции могут происходить значительные изменения возрастной структуры. В таких случаях «включаются механизмы», автоматически возвращающие популяцию к некоторому нормальному для данной популяции возрастному распределению. Причем популяции, включающие в себя много возрастных групп, в меньшей степени подвержены воздействию факторов, влияющих на размножение в конкретном году.
Даже крайне неблагоприятные условия, вызывающие полную гибель приплода в тот или иной год, не становятся катастрофическими для популяции сложной структуры, так как родительские пары участвуют в воспроизводстве многократно. Тем не менее следы изменений условий жизни сохраняются в облике сложных популяций гораздо дольше, чем у простых.
Для роста численности популяций большое значение имеет соотношение особей по полу. Генетический механизм обеспечивает примерно равное соотношение особей разного пола при рождении. Однако исходное соотношение вскоре нарушается в результате различий
физиологических, поведенческих и экологических реакций самцов и самок, вызывающих неравномерную смертность.
Анализ возрастной и половой структуры популяции позволяет прогнозировать ее численность на ряд ближайших поколений и лет. Этим пользуются для оценки возможности промысла рыбы, в охотничьих хозяйствах и прочих подобных случаях.
Пространственная и этологическая структуры популяций
Пространственная структура. Каждая популяция занимает пространство, обеспечивающее условия жизни для ограниченного числа особей. При изучении пространственной структуры различают случайное, равномерное и неравномерное (групповое) распределения особей на территории (в пространстве).
Случайное распределение в природе встречается редко; оно наблюдается в случаях, когда среда очень однородна, а организмы не стремятся объединиться в группы. Равномерное распределение, бывает там, где между особями очень сильна конкуренция или существует антагонизм. Наиболее часто наблюдается неравномерное (групповое) распределение — образование различных скоплений.
Активность особей, пар и семейных групп у позвоночных и высших беспозвоночных обычно ограничена определенной зоной, называемой индивидуальным (или семейным) участком территории. У высших животных внутрипопуляционное распределение регулируется системой инстинктов. Им свойственно особое территориальное поведение — реакция на местонахождение других членов популяции.
В зависимости от характера использования пространства подвижных животных подразделяют на оседлых и кочевых. Оседлые животные в течение всей или большей части жизни используют довольно ограниченный участок среды. Им присущи инстинкты привязанности к своему участку, регулярное возвращение к месту размножения после длительных и дальних миграций.
Кочевые животные совершают постоянные передвижения в пространстве, так как они зависят от запаса корма на данной территории. Кочевой образ жизни характерен преимущественно для стад и стай (кочующие группы многих рыб во время нагульных миграций, стада слонов, зебр, антилоп, северных оленей и т. п.) Перемещение кочующих видов по площади обычно происходит за сроки, достаточные для восстановления кормовых ресурсов на пастбищных участках. Масштабы и длительность таких миграций определяют обилие пищи и численность стада.
Этологическая (поведенческая) структура
Она отражает разнообразные формы совместного существования особей в популяциях. Одиночный образ жизни следует выделить в первую очередь, хотя полностью одиночного существования организмов в природе нет, так как в этом случае было бы невозможно размножение. Семейный образ жизни — усиливаются связи между родителями и потомством, начинает заметно проявляться территориальное поведение животных. Путем различных сигналов, маркировки, угроз и тому подобного обеспечивается владение участком, достаточным для выкармливания потомства.
Стая — временное объединение животных, проявляющих биологически полезную организованность действий (для защиты от врагов, добычи пищи, миграции и т. п.). Наиболее широко стайность распространена среди рыб и птиц, хотя встречается и у млекопитающих (например, у собак).
Стадо — длительное или постоянное объединение животных, в котором осуществляются все основные функции жизни вида: добывание корма, защита от хищников, миграция, размножение, воспитание молодняка.
Основу группового поведения в стадах составляют взаимоотношения доминирования. Характерно наличие временного или относительно постоянного лидера, которому подражают особи стада.
Активное руководство стадом (специальные сигналы или угрозы) осуществляют вожаки. Иерархически организованному стаду свойственен закономерный порядок перемещения, определенные позиции при защите от врагов, расположение на местах отдыха и др.
Колония — это групповое поселение оседлых животных на длительное время или на период размножения. По сложности взаимоотношений между особями колонии очень разнообразны, наиболее сложные отношения складываются в поселениях для общественных насекомых (термитов, муравьев, пчел, ос и др.), возникающие на основе сильно разросшейся семьи. Члены колоний постоянно обмениваются информацией друг с другом.
Динамика популяций
Динамика популяций — это процессы изменения ее основных биологических показателей (численности, биомассы, структуры) во времени в зависимости от экологических факторов. Жизнь популяции проявляется в ее динамике — одном из наиболее значимых биологических и экологических явлений.
Кривые выживания
Кривая выживания представляет собой график зависимости от времени числа выживших на данный момент особей для некоторой начальной группы новорожденных. Каждому биологическому виду свойственна своя характерная кривая выживания. При построении графика по оси абсцисс откладывают относительный возраст, а по оси ординат — абсолютное число выживших особей или их процент от исходного числа.
Все животные и растения подвержены старению, которое проявляется в снижении жизненной активности с возрастом после периода зрелости. Непосредственные причины смерти могут быть разными, но в основе их лежит уменьшение сопротивляемости организма к неблагоприятным факторам, например болезням.
Причиной смерти преимущественно служит случай, и особи гибнут до начала заметного старения. Кривая, подобная этой, была получена, например, для «популяции» стеклянных (легко бьющихся) стаканов в кафетерии. В природе к таким видам относятся, например, устрицы, дающие огромное потомство, а также растения, размножающиеся благодаря рассеиванию большого числа семян.
К промежуточному типу относят кривые выживания таких видов, для которых смертность мало меняется с возрастом. В природе существует много видов птиц, ящериц, мелких млекопитающих и других организмов, имеющих выживания, которые отличаются от прямолинейной диагонали некоторой выпуклостью (вогнутостью) или волнообразностью.
Форма выживания часто меняется при изменении плотности популяции. С увеличением плотности кривая становится более вогнутой, т. е. при увеличении численности организмов их смертность возрастает.
Рост популяции
Если при незначительной эмиграции и иммиграции рождаемость превышает смертность, то популяция будет расти. Рост популяции является непрерывным процессом, если в ней существуют все возрастные группы. Скорость роста популяции при отсутствии каких либо экологических ограничений.
Такая модель основывается на допущении, что рост популяции не зависит от ее плотности. Считают, что почти любой вид теоретически способен увеличить свою численность до заселения всей Земли при достатке пищи, воды, пространства, постоянстве условий среды и отсутствии хищников. Эта идея была выдвинута еще на рубеже XVIII и XIX вв. английским экономистом Томасом Р. Мальтусом, основоположником теории мальтузианства.
образная модель роста популяции.
Иное развитие получает ситуация при ограниченности пищевых ресурсов либо при скоплении токсичных продуктов (отходов) метаболизма. Первоначальный экспоненциальный рост в исходных благоприятных условиях со временем продолжаться не может и постепенно замедляется. Плотность популяции регулирует истощение пищевых ресурсов, накопление токсикантов и поэтому влияет на рост численности. С увеличением плотности скорость роста популяции постепенно снижается до нуля, и кривая выходит на некоторый стабильный уровень (график образует плато). Кривая такого роста имеет Sобразную форму, и поэтому соответствующая модель развития событий называется S-образной.
Она характерна, например, для дрожжей, фактором, ограничивающим их рост, является накопление спирта, а также для водорослей, самозатеняющих друг друга. В обоих случаях численность популяции не достигает уровня, на котором начинает сказываться нехватка элементов питания (биогенов).
На рост численности, в которой значительную (возможно, даже главную) роль играет пространство, также влияет перенаселенность.
Миграция или расселение, так же как и внезапное снижение скорости размножения, могут способствовать уменьшению численности популяции. Расселение может быть связано с определенной стадией жизненного цикла, например с образованием семян.
Применительно к условиям реальной природной среды принято использовать понятия биотический потенциал — совокупность всех экологических факторов, способствующих увеличению численности популяции, или видовая способность к размножению при отсутствии ограничений со стороны среды, а также сопротивление среды — сочетание факторов, ограничивающих рост (лимитирующих факторов).
Любые изменения популяции есть результат нарушения равновесия между ее биотическим потенциалом и сопротивлением окружающей среды.
Колебания численности популяции
По достижении заключительной фазы роста размеры популяции продолжают колебаться от поколения к поколению вокруг некоторой более или менее постоянной величины. При этом численность одних видов изменяется нерегулярно с большой амплитудой колебаний (насекомыевредители, сорняки), колебания численности других (например, мелких млекопитающих) имеют относительно постоянный период, а в популяциях третьих видов численность колеблется от года к году незначительно (долгоживущие крупные позвоночные и древесные растения).
В природе в основном встречаются три вида кривых изменения численности популяции: относительно стабильный, скачкообразный и циклический .
Виды, у которых численность из года в год находится на уровне поддерживающей емкости среды, имеют достаточно стабильные популяции. Такое постоянство характерно для многих видов дикой природы и встречается, например, в нетронутых тропических влажных лесах, где среднегодовое количество осадков и температура изменяются день ото дня и из года в год крайне мало.
У других видов колебания численности популяций носят правильный циклический характер.
Ряд видов, таких, как енот, в основном имеют достаточно стабильные популяции, однако время от времени их численность резко возрастает (подскакивает) до наивысшего значения, а затем резко падает до некоторого низкого, но относительно стабильного уровня. Эти виды относят к популяциям со скачкообразным ростом численности Внезапное увеличение численности происходит при временном повышении емкости среды для данной популяции и может быть связано с улучшением климатических условий (факторов) и питания или резким уменьшением численности хищников (включая охотников). После превышения новой, более высокой емкости среды в популяции возрастает смертность и ее размеры резко сокращаются.
На протяжении истории в разных странах не раз наблюдались случаи краха популяций человека, например, в Ирландии в 1845 г., когда в результате заражения грибком погиб весь урожай картофеля. Поскольку рацион питания ирландцев сильно зависел от картофеля, к 1900 г. половина восьмимиллионного населения Ирландии умерла от голода или эмигрировала в другие страны.
Тем не менее численность человечества на Земле в целом и во многих регионах в частности продолжает расти. Люди путем технологических, социальных и культурных перемен неоднократно увеличивали для себя поддерживающую емкость планеты. По сути, они смогли изменить свою экологическую нишу за счет увеличения производства продуктов питания, борьбы с болезнями и использования больших количеств энергетических и материальных ресурсов, чтобы сделать обычно непригодные для жизни районы Земли обитаемыми.
Популяции синантропных видов
В предыдущих разделах рассмотрены природные популяции, находящиеся в естественных местообитаниях. Однако вследствие хозяйственной деятельности человека образуются природноантропогенные популяции, тесно связанные, например, с сельским хозяйством. Многие насекомые, мышевидные грызуны и прочие виды находят здесь экологическую нишу, адаптируя свою структуру и динамику численности к той или иной системе хозяйствования.
С эволюцией человеческого общества и образованием стабильных поселений также возникли синантропные виды, популяции которых обитают в жилищах и в местах скопления отходов жизнедеятельности людей. Они могут быть «вредителями», «паразитами», переносчиками возбудителей болезней, выступая в качестве опасных экологических (биотических) факторов для человека.
В пещеру древнего человека перешли на жительство многие насекомые, осваивая скопления различных отходов. Однако человечество постепенно развивало культуру быта и улучшало санитарногигиеническую обстановку в своих поселениях, одновременно создавая разнообразные методы уничтожения нежелательных « спутников».