

Белки / Сложные белки
.docxНуклеопротеины. Небелковая часть представлена НК: ДНК и РНК. НК участвуют в хранении и передаче наследственной информации о 1 структуре белков. Белковая часть нуклеопротеинов - протамины и гистоны.
Хромопротеины. Относятся гемоглобин, миоглобин, церулоплазмин. Гемоглобин состоит из глобина и небелковой части - гема, в составе которого имеется атом двухвалентного железа и окрашен в красный цвет. Белок крови церулоплазмин, содержащий медь, окрашен в голубой цвет.
Липопротеины являются компонентами клеточных мембран, содержатся в миелиновых оболочках нервных клеток, а также в синаптических мембранах и внутренних мембранах митохондрий.
Гликопротеины входят в состав клеточных мембран, участвуют в транспорте веществ, в процессах свертывания крови, иммунитета (иммуноглобулины), образуют матрикс межклеточного пространства.
Фосфопротеины - белки, играющие роль в питании молодых организмов. Это основные белки молока - казеин, яичного желтка – вителлин и фосвитин, икры рыб – ихтулин.
Металлопротеины - алкогольдегидрогеназа, содержащая цинк. К медьсодержащим белкам относятся цитохромоксидаза, церулоплазмин, к железосодержащим – лактоферрин молока, трансферрин.
* * *
К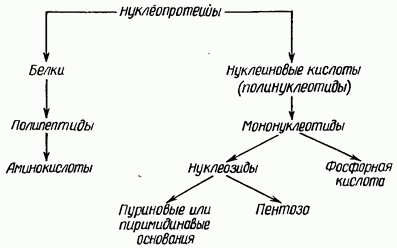 ислотный
гидролиз в присутствии серной кислоты.
При непродолжительном нуклеопротеиды
распадаются на белок и НК. При
продолжительном - полный распад
нуклеопротеидов.
ислотный
гидролиз в присутствии серной кислоты.
При непродолжительном нуклеопротеиды
распадаются на белок и НК. При
продолжительном - полный распад
нуклеопротеидов.
При гидролизе мононуклеотидов выделяются пуриновые или пиримидиновые основания, углевод и фосфорная кислота.
В составе белковых компонентов очень много положительно заряженных аминокислот – аргинина и лизина, поэтому их можно отнести к поликатионам (гистоны).
***
Молекулы НК заряжены отрицательно, образуют с + заряженными белковыми компонентами ионные связи.
НК – линейные (циклические) гетерополимеры, их мономерами являются мононуклеотиды.
Мононуклеотид состоит из 3 частей: 1) азотистого основания (пуринового или пиримидинового), 2) пентозы (рибозы, дезоксирибозы) – вместе с азотистым основанием они составляют нуклеозид, 3) остатка фосфорной кислоты.
1. ДНК: хранение генетической информации.
2. РНК:
а) хранение ген. информации (информосомы, некоторые РНК-вирусы);
б) реализация ген. информации: и-РНК, т-РНК, р-РНК.
в) каталитическая функция: способны катализировать реакции гидролиза 3',5'-фосфодиэфирной связи в самой молекуле РНК, способны одним участком катализировать гидролиз фосфодиэфирных связей другого участка своей молекулы «самосплайсинг».
 ***
***
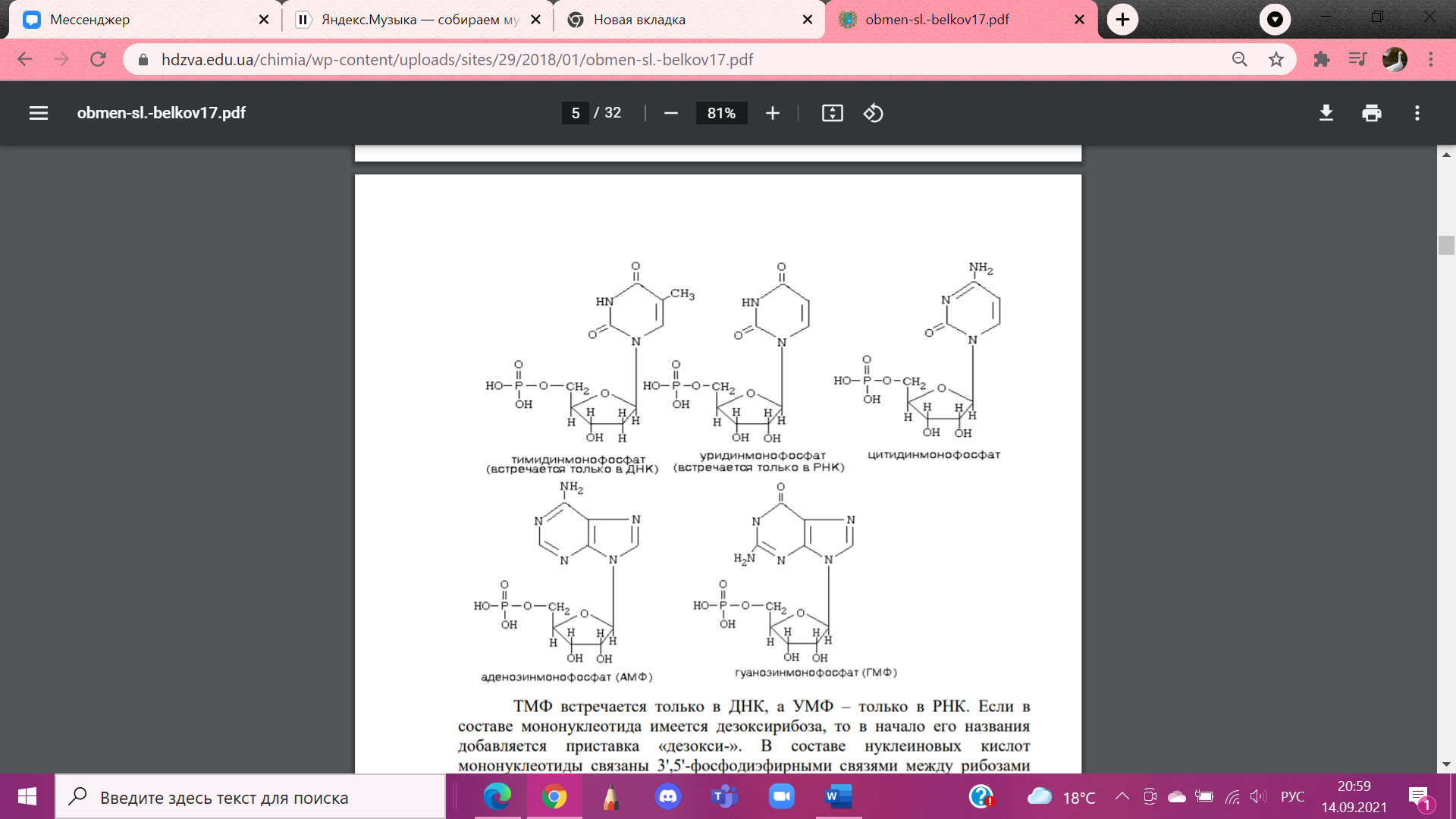
 В
составе НК мононуклеотиды связаны
3',5'-фосфодиэфирными связями между
рибозами соседних мононуклеотидов
через остаток фосфорной кислоты. Если
молекула НК не является циклической,
концы ее различны. Один из них обозначается
как 3'-конец, а другой – 5'-конец (начальный).
В
составе НК мононуклеотиды связаны
3',5'-фосфодиэфирными связями между
рибозами соседних мононуклеотидов
через остаток фосфорной кислоты. Если
молекула НК не является циклической,
концы ее различны. Один из них обозначается
как 3'-конец, а другой – 5'-конец (начальный).
***
1 структура - цепочки из нуклеотидов, соединяющихся через остаток фосфорной кислоты (фосфодиэфирная связь).
2 структура - 2 цепи НК, соединённые водородными связями (соединяются по типу «голова-хвост» (3' к 5'), по принципу комплементарности (азотистые основания находятся внутри этой структуры)).
3 структура (спираль) за счет радикалов азотистых оснований (образуются водородные дополнительные связи, они сворачивают структуру, обуславливая прочность).
4 структура - комплексы гистонов и нитей хроматина.
Согласно модели Крика - Уотсона, ДНК - двойная спираль, состоящая из 2 цепей дезоксирибозофосфата, соединённых парами оснований аналогично ступенькам лестницы. Посредством водородных связей аденин соединяется с тимином, а гуанин — с цитозином. С помощью этой модели можно было проследить репликацию самой молекулы ДНК. По Уотсону и Крику, 2 части молекулы ДНК отделяются друг от друга в местах водородных связей. Из каждой половины молекулы синтезируется новая ДНК. Последовательность оснований функционирует как матрица для новых ДНК.
Кол-во аденина равно количеству тимина, а гуанина - цитозину.
Количество пуринов равно количеству пиримидинов: А+Г=Т+Ц.
К-во оснований с аминогруппами в положении 6 равно количеству оснований с кетогруппами в положении 6: А+Ц=Т+Г.
 ***
***
Циклические нуклеотиды, к которым относятся 3'-,5'-циклоаденозин- и 3'-,5'-циклогуанозинмонофосфат, являются вторичными посредниками в действии полипептидных гормонов на метаболизм клетки
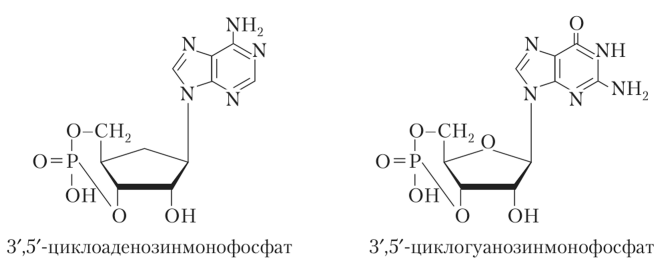
Они образуются из нуклеозидтрифосфатов путем формирования фосфодиэфирной связи между 3'- и 5'-атомами углерода пентозного цикла под действием ферментов аденилат- и гуанилатциклаз и вместе с соответствующими протеинкиназами участвуют в фосфорилировании внутриклеточных белков (ферментов), изменяя их конформацию и активность.