

ВИРУС ИММУНОДЕФИЦИТА ЧЕЛОВЕКА
Характеристика ретровирусов
•Однонитевые РНК-содержащие вирусы (РНК +)
•Имеется обратная транскриптаза (РНК- зависимая ДНК-полимераза)
•Имеются протеины: группового АГ (gag), полимеразный протеин (pol) и белки оболочки (env)
Вирус иммунодефицита человека (ВИЧ, или HIV – от анг. Human Immuodeficiency Virus) -
Вызывает ВИЧ-инфекцию, заканчивающуюся развитием синдрома приобретенного иммунодефицита (СПИД, или от анг. AIDS - Acquired Immuodeficiency Syndrome). ВИЧ характеризуется преимущественным поражением иммунной системы, длительным течением, полиморфностью клинических проявлений, высокой летальностью, передачей в естественных условиях от больного человека здоровому и склонностью к быстрому эпидемическому распространению. Типичный антропоноз
Относится к семейству Retroviridae, роду Lentivirus


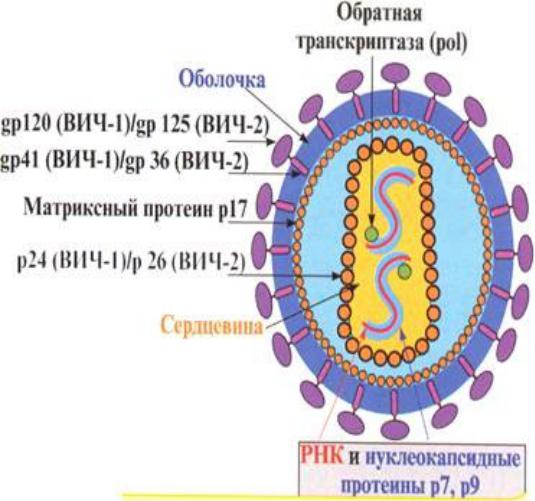
Строение вируса ВИЧ
•Вирусная частица имеет сферическую форму
•Оболочка вируса состоит из двойного слоя липидов, пронизанного гликопротеинами
•Липидная оболочка происходит из плазматической мембраны клетки хозяина, в которой репродуцируется вирус
•Гликопротеиновая молекула 160кДа, состоит из 2 субъединиц: gp 120 (молекулярная масса 120 кДа), находящейся на поверхности вириона, и gp 41 (молекулярная масса 41 кДа), пронизывающей его липидную оболочку
•Сердцевина вируса состоит из капсидных белков р24, р25, ряда матриксных белков (р6, р7) и белков протеазы (р10, р11)
•Геном представлен двумя нитями РНК
•Имеют обратную транскриптазу (ревертазу)
•Геном состоит из 3 основных структурных генов (gag, pol, env) и 7 регуляторных и функциональных генов (tat, rev, nef, vif, vpr, vpu, vpx)
•Ген gag (group antigen) кодирует матриксные, капсидные, нуклкеокапсидные, белки и белки протеазы
•Ген pol (polymerase) кодирует обратную транскриптазу
•Ген env (envelope) кодирует поверхностный белок gp 12 и трансмембранный gp41
•Функциональные гены выполняют регуляторные функции 9reg, tat, nef) и обеспечивает осуществление процессов репродукции и участие вируса в инфекционном процессе 9vif, vpu, vpr, vpx)
•Проведенные детальные исследования свойств ВИЧ-2 показали, что при определенном сходстве с ВИЧ-1, он отличается от последнего по антигенной структуре и по последовательности оснований в нуклеиновых кислотах. ВИЧ-2 более близок по своим свойствам (в том числе по антигенной структуре и составу генетического материала) к вирусу иммунодефицита обезьян (SIV/ВИО), чем к ВИЧ-1
•ВИЧ-1 и ВИЧ-2 инфицируют те же популяции клеток, связываются с теми же CD4-рецепторами. Проведен анализ полного генома ВИЧ-2. В целом его организация сходна с таковой ВИЧ-1. В дополнение к классическим ретровирусным генам gag, pol и env оба вируса содержат гены vif, vpr, tat, nef и rev
•Одним из наиболее существенных различий между вирусами является наличие у ВИЧ-1 гена vpu, не обнаруживаемого у ВИЧ-2. В тоже время ВИЧ-2 содержит ген vpx, которого нет у ВИЧ-1. Продуктом гена vpu является белок с молекулярной массой 14-16 кД, а гена vpx - белок с молекулярной массой 12-14 кД. Продукт гена vpu весьма важен для созревания и высвобождения вируса из клетки. Гены tat, ref и nef необходимы для вирусной репликации и экспрессии вирусного генома. Роль генов vif и vpr пока неясна. Полагают, что vif может играть роль в посттрансляционной модификации вирусных белков.
•Наиболее консервативными участками геномов ВИЧ-1 и ВИЧ-2 являются гены gag и pol. Они соответственно имеют 57 и 59% гомологии. Менее консервативен участок, кодирующий оболочечные белки, особенно содержащиеся в наружной части оболочки вируса (37% идентичных аминокислот). Было установлено, что большинство цистеиновых остатков в ВИЧ-1 и ВИЧ-2 стабильно, это свидетельствует о том, что третичная структура белка может иметь важное функциональное значение. Трансмембранная часть оболочечного белка - gp41- более стабильна, чем наружная - gp120. Важную роль в цитопатогенности ВИЧ отводят С-концу трансмембранного белка. N-терминальный конец этого белка является уникальным регионом, специфичным для каждого из ВИЧ. На основании этой уникальности можно осуществить дифференциацию ВИЧ-1 и ВИЧ-2. ВИЧ-2 отличается от ВИЧ-1 по оболочечным гликопротеинам и в меньшей степени - по сердцевинным антигенам. Сыворотки большинства людей, инфицированных ВИЧ-2, не реагируют с оболочечными антигенами ВИЧ-1, но дают перекрестные реакции с антигенами, кодируемыми геном gag.
•Имеются различия в молекулярной массе белков ВИЧ-1 и ВИЧ-2. В частности, наружный оболочечный гликопротеин имеет молекулярную массу 120 кД, у ВИЧ-2 этот белок имеет мол. массу 140 кД. Трансмембранный белок у ВИЧ-2 - 36 кД, а у ВИЧ-1 - 41кД Белки кодируемые геном gag у ВИЧ-1 имеют мол. массу 24 и 17 кД, а у ВИЧ-2 - 26 и 15 кД.

Жизненный цикл ВИЧ
1.Адсорбция и проникновение вируса путем эндоцитоза в клетку
2.Высвобождение вирусной РНК, на которой синтезируется комплементарная минус-нить ДНК. С нее копируется плюс-нить с образованием двойной кольцевой ДНК. Комплементарная ДНК интегрирует с хромосомной ДНК клетки
3.Рекомбинантная ДНК-провирус служит основой синтеза геномной РНК вируса и иРНК, которые обеспечивают синтез компонентов и сборку вирионов
4.Вирионы выходят из клетки почкованием: сердцевина вируса «одевается» в модифицировванную плазмолемму клетки
Полный жизненный цикл вируса реализуется за 1-2 сут, в сутки формируется до одного миллиарда вирусных частиц
Проникновение в клетку и обратная транскрипция
Вирусный гликопротеин gp120 прочно связывает рецептор CD4. В результате такого взаимодействия gp120 претерпевает конформационные изменения, которые позволяют ему также связать молекулу корецептораCXCR4 или CCR5 (экспрессируемых на поверхности Т- лимфоцитов, макрофагов, дендритных клеток и микроглии). В зависимости от способности связывать эти корецепторы, ВИЧ классифицируют на R5-тропные (связывают только корецептор CCR5), X4-тропные (связывают только корецептор CXCR4) и R5X4-тропные (могут взаимодействовать с обоими корецепторами) варианты. При заражении, в основном, передаются R5-тропные и R5X4-тропные варианты. Препараты, блокирующие корецепторы, могут быть эффективны против ВИЧ.
После описанных событий вирусный белок gp41 проникает в мембрану клетки и подвергается значительным конформационным изменениям, вследствие которых мембрана клетки и мембрана вириона ВИЧ сближаются друг с другом и затем сливаются. Вирусный белок gp41 очень важен для слияния мембран, поэтому его рассматривают в качестве мишени для разработки противовирусных препаратов.
После слияния мембран содержимое вириона проникает внутрь клетки. Внутри клетки вирусная РНК высвобождается из капсида. Затем под действием обратной транскриптазы происходит обратная транскрипция — процесс синтеза ДНК на основании информации в одноцепочечной геномной РНК вируса. Большая часть лекарственных препаратов, одобренных для применения при ВИЧ-инфекции, направлена на нарушение работы обратной транскриптазы
Транспорт вирусной ДНК в ядро и интеграция в геном
После завершения обратной транскрипции в CD4+-лимфоците вирусный геном представлен невстроенной ДНК. Для встраивания вирусной ДНК в геном клетки- хозяина и для образования новых вирусов необходима активация Т-лимфоцитов. Активация CD4+-лимфоцитов происходит при их контакте с антигенпредставляющими клетками в лимфоидной ткани. Наличие вирусов на поверхности фолликулярных дендритных клеток и присутствие провоспалительных цитокинов (ИЛ-1, ИЛ-6, ФНОα) способствуют размножению ВИЧ в инфицированных клетках. Именно поэтому лимфоидная ткань служит самой благоприятной средой для репликации ВИЧ
Синтезированная вирусная ДНК транспортируется внутрь ядра клетки в составе пре-интеграционного комплекса, в который также входят белки ВИЧ p17/MA, Nef и интеграза. Далее вирусная ДНК встраивается в хромосомы активированного T- лимфоцита под действием интегразы. Несколько препаратов, ингибирующих интегразу, широко используются в современной комплексной антиретровирусной терапии. Вирусная ДНК, встроившаяся в хромосому клетки, называется провирусом